
Glomerella cingulata
Glomerella cingulata est un champignon parasite qui cause des maladies sur différents fruits et légumes sous le nom d'anthracnose avec une aire de répartition mondiale de la maladie. Des plantes cultivées comme l'herbe de la Saint-Jean (Hypericum perforatum) sont touchées. Il cause aussi la maladie des feuilles de l'Hevea brasiliensis.
 Glomerella cingulata sur Umbellularia californica (Bitter rot)
Glomerella cingulata sur Umbellularia californica (Bitter rot)
Glomerella cingulata est le stade sexué (téléomorphe) alors que le stade asexué (anamorphe) est désigné par Colletotrichum gloeosporioides.
Réaction végétale de défense
La daniélone est un phénol naturel, une phytoalexine produite par la papaye Carica papaya pour combattre Colletotrichum gloeosporioides.
Glomerella cingulata a été associé à des maladies chez un large éventail de plantes hôtes. On le trouve couramment comme agent pathogène et endophyte dans les cultures et les arbres tropicaux, ainsi que dans l'infection des semis d'arbres dans les forêts tempérées. G. cingulata peut infecter les fruits, les fleurs, les feuilles et d'autres tissus non lignifiés. La phylogénie de ce groupe n'est pas bien résolue; sur la base de phylogénies multi-gènes, au moins 22 taxons sont connus dans le complexe d'espèces. La plupart des membres de ce groupe sont mal compris en termes de biologie, de pathogénicité et de préférence pour l’hôte. Certaines des souches sont à l'étude pour leur activité catalytique sur des composés végétaux complexes.
Les caractéristiques culturelles ont été étudiées afin de distinguer les isolats de Glomerella cingulata et de Colletotrichum spp. associée à la tache foliaire à Glomerella et à la pourriture amère des pommes chez celles qui ne causent que la pourriture amère. Le taux de croissance, la réponse à la température et la sensibilité au bénomyl de 27 isolats de Glomerella cingulata , de 12 isolats de Colletotrichum gloeosporioides et de 7 isolats de C. acutatum , récoltés dans des vergers situés aux États-Unis et au Brésil et caractérisés précédemment en fonction de la morphologie, de la végétation compatibilité et les haplotypes d’ADN mitochondrial (ADNmt) ont été déterminés. Ces isolats représentent la diversité génétique et moléculaire au sein des isolats de C. gloeosporioides, C. acutatum et G. cingulata de pommes trouvés dans une étude précédente. Une croissance plus lente, une température de croissance optimale inférieure et une sensibilité moindre au bénomyl distinguent les isolats de C. acutatum des isolats de G. cingulata et de C. gloeosporioides. Cependant, le taux de croissance et la sensibilité au bénomyl ne sont pas utiles pour distinguer G. cingulata de C. gloeosporioides ni pour différencier les isolats de G. cingulata qui causent la tache des feuilles et la pourriture amère de ceux qui ne provoquent que la pourriture amère.
Introduction
|
Fig 1. Maladie de la pourriture amère des pommes ( Colletotrichum gloeosporioides , C. acutatum et Glomerella cingulata ). |

La pourriture amère est une maladie fréquente du fruit de la pomme dans pratiquement tous les pays où elle est cultivée commercialement. Dans les régions à croissance humide et tempérée, il est considéré comme l'une des maladies les plus importantes et peut causer des pertes de récolte pouvant atteindre 50% (16). Trois taxons, Colletotrichum gloeosporioides (Penz.) Penz. & Sacc., C. acutatum JH Simmonds et Glomerella cingulata (Stonem.) Spauld. Et Schrenk, sont associés à la pourriture amère des pommes (16) (Fig. 1). En 1988, Leite et al. (13) ont décrit une nouvelle maladie des taches du pommier sur les cultivars Gala et Golden Delicious dans l'État de Paraná au Brésil et l'ont associée à des isolats homothalliques de C. gloeosporioides, désignés par G. cingulata (14).Ces isolats produisent des périthèces fertiles et sont souvent appelés isolats périthéciaux (15). Il s'agissait du premier signalement de Colletotrichum spp.provoquant une tache sur les pommes. La maladie, appelée Glomerella leaf spot (GLS), se caractérise par de petites taches nécrotiques irrégulières sur les feuilles (Fig. 2). Sa gravité a augmenté et est devenue une préoccupation majeure pour les pomiculteurs brésiliens, car l'un des cultivars les plus cultivés, Gala, est très sensible à la maladie. Dans des conditions favorables, GLS peut entraîner une défoliation à 75% (Fig. 3) du fait des récoltes, affaiblir les pommiers et réduire le rendement (4, 13,17). De plus, GLS a été observé sur d'autres cultivars importants sur le plan commercial cultivés au Brésil, tels que Granny Smith et Pink Lady (Dr. Rosa Maria Sanhueza, communication personnelle, 2002). GLS a été signalé pour la première fois aux États-Unis en 1998 en tant que tache grave des feuilles sur le cv. Des pommes Gala dans deux vergers de l’est du Tennessee (9) ont été signalées par la suite en Géorgie et en Caroline du Nord. Bien que GLS ait été signalé pour la première fois aux États-Unis en 1998, des expériences menées en Géorgie à la fin des années 1960 (19) avec un isolat de G. cingulata suggèrent que des souches du champignon capables de causer la tache des feuilles étaient présentes beaucoup plus tôt.
|
|
|
|||
|
|
 |
|

|
|
Fig. 2. Tache de la feuille de Glomerella (Glomerella cingulata) sur les pommes, caractérisée par une tache de la feuille irrégulière. |
Fig. 3. Défoliation dans les pommiers causée par GLS. Photo gracieuseté de Dr. Rosa Maria Sanhueza. |
Bien que GLS et la pourriture amère soient associés au même champignon, des différences de morphologie, de caractéristiques de culture et de pouvoir pathogène ont été observées entre les isolats de l'agent pathogène obtenus à partir de fruits ou de feuilles (13,19). Ces différences, associées à la variabilité génétique et moléculaire considérable au sein des isolats de C. gloeosporioides et G. cingulata obtenus à partir de fruits présentant des symptômes de pourriture amère (3,15) et au sein de populations de C. gloeosporioides et de G. cingulata d’autres cultures (5,6, 7, 11, 20, 21) suggèrent que la diversité de ces agents pathogènes présents sur les pommes est élevée. Une étude récente portant sur la caractérisation morphologique, génétique et moléculaire d'isolats de G. cingulata et C. acutatum et de C. gloeosporioides obtenus à partir de pommes récoltées à différents endroits aux États-Unis et au Brésil a révélé une grande diversité au sein de chaque espèce (8). . Douze types morphologiques, 16 GCV, 12 haplotypes d'ADNmt et 7 groupes phylogénétiques, résumés dans le tableau 1, ont été trouvés parmi les isolats. En outre, des tests de pathogénicité foliaire ont montré que seuls les isolats de G. cingulata présentant des GCV spécifiques et des haplotypes étaient pathogènes pour les feuilles (tableau 1).
Tableau 1. Caractérisation et pathogénicité des isolats de G. cingulata, C. gloeosporioides et C. acutatum en fonction de groupes morphologiques, génétiques et moléculaires trouvés parmi les isolats collectés dans des vergers de pommiers situés aux États-Unis et au Brésil x .
|
Espèce |
Séquence |
ADNmt |
VCG |
Morpho- |
Feuille |
Emplacement |
|
G. |
Groupe 1 |
G1 |
1 |
SP1 |
+ |
NOUS |
|
1 |
SP1 |
+ |
NOUS |
|||
|
2 |
SP1, SP2 |
- |
NOUS |
|||
|
3 |
SP1 |
- |
NOUS |
|||
|
G1.1 |
1 |
SP1 |
+ |
NOUS |
||
|
1 |
SP1 |
- |
NOUS |
|||
|
2 |
SP2 |
- |
NOUS |
|||
|
G2 |
2 |
SP2 |
- |
NOUS |
||
|
G2.1 |
2 |
SP2 |
- |
NOUS |
||
|
Groupe 2 |
G3 |
4 |
SP1 |
+ |
Brésil |
|
|
5 |
SP1 |
+ |
Brésil |
|||
|
G4 |
5 |
SP1 |
+ |
Brésil |
||
|
Groupe 3 |
A3 |
6 |
CP |
- |
NOUS |
|
|
A3.1 |
6 |
SP3 |
- |
NOUS |
||
|
C. gloeo- |
Groupe 4 |
B2 |
7 |
SS2 |
- |
NOUS |
|
12 |
SS5 |
- |
NOUS |
|||
|
Groupe 5 |
B2 |
n / a |
- |
NOUS |
||
|
B3 |
8 |
SS3 |
- |
NOUS |
||
|
9 |
SS1 |
- |
NOUS |
|||
|
Groupe 6 |
B2 |
11 |
SS3 |
- |
NOUS |
|
|
B3 |
11 |
SS3 |
- |
NOUS |
||
|
Groupe 7 |
B2 |
dix |
SS1 |
- |
NOUS |
|
|
C. |
n / a |
C1 |
15 |
SSC |
- |
NOUS |
|
n / a |
16 |
SSNC |
n / a |
NOUS |
||
|
n / a |
13 |
SSNC (O) |
n / a |
Brésil |
||
|
D1 |
14 |
SSNC (O) |
- |
Brésil |
x Analyse de séquence (groupes basés sur les arbres phylogénétiques de maximum de vraisemblance et de parcimonie maximale, basés sur la séquence d'un intron de 200 pb du gène nucléaire GDPH), d'haplotypes d'ADN mt (groupes RFLP d'ADN génomique digérés avec Msp I), de VCG (groupes de compatibilité végétative) , les types morphologiques (basés sur la couleur des colonies, la forme des conidies, la capacité de produire des périthèces en culture et la distribution des acervuli et des périthèces en culture) et la pathogénicité (déterminée par l'inoculation d'arbres de cv. Gala cultivés en serre) étude précédente (8). '+' indique pathogène; '-' indique non pathogène.n / a = données non disponibles.
Des caractéristiques culturelles ont également été utilisées pour distinguer les isolats de C. gloeosporioides, G. cingulata et C. acutatum. Les isolats de G. cingulata et de C. gloeosporioides se caractérisent par un taux de croissance plus rapide (10, 12, 14,18) et une plus grande sensibilité au bénomyl (1, 2, 12,18).De plus, les isolats de C. acutatum ont montré des températures de croissance optimales inférieures à celles des isolats de C. gloeosporioides (1). Les outils génétiques et moléculaires peuvent prendre du temps et coûter cher. Par conséquent, l'utilisation de caractéristiques culturelles peut fournir un moyen plus rapide et moins coûteux de différencier les isolats de G. cingulata associés à GLS.
L’objectif de cette étude était d’utiliser le taux de croissance, la réponse à la température et la sensibilité au bénomyl pour déterminer s’il existe des différences entre les haplotypes d’ADNmt de C. gloeosporioides, C. acutatum et G. cingulata et, s’il existe, différencier peut être utilisé pour distinguer les isolats de G. cingulata associés à GLS de ceux ne causant que la pourriture amère.
Caractérisation culturelle des isolats
Vingt-sept isolats de G. cingulata (14 de fruits et 13 de feuilles), 12 isolats de fruits de C. gloeosporioides et 7 isolats de C. acutatum (quatre de fruits et trois de feuilles) ont été sélectionnés dans la collection d'isolats monosporique. , précédemment caractérisées sur la base de la morphologie, de la compatibilité végétative et des haplotypes RFLP de mDADNA (8) (Tableau 2) afin de déterminer le taux de croissance, la température de croissance optimale et la sensibilité des isolats au bénomyl. Les isolats représentaient les haplotypes A3, B2, B3, C1, D1, G1, G2, G3 et G4. Des isolats de taches sur les feuilles de G. cingulata se trouvent dans les haplotypes G1, G3 et G4. Le taux de croissance (en mm / jour) des isolats de G. cingulata, C. gloeosporioides et C. acutatum a été déterminé en mesurant le diamètre de la colonie des isolats toutes les 24 h pendant 6 jours à 25 ° C avec une lumière constante. Des boîtes de Pétri contenant environ 15 ml de PDA ont été ensemencées avec des bouchons de 5 mm de diamètre de chaque isolât, obtenus à partir des marges de cultures âgées de 4 jours. Trois boîtes de Pétri ont été utilisées pour chaque isolat et l'expérience a été répétée une fois. La sensibilité au bénomyl a été déterminée en modifiant des boîtes de Pétri contenant 15 ml de PDA à chacune des concentrations suivantes: 0, 0,01, 0,1, 1 et 10 µg / ml. Trois boîtes de Pétri ont été utilisées par isolat et deux essais ont été réalisés.
Tableau 2. Isolats de G. cingulata, C. gloeosporioides et C. acutatum examinés pour déterminer leur taux de croissance, leur réponse à la température et leur sensibilité au bénomyl x .
|
Espèce |
Isoler |
Géo- |
La source |
ADN mt |
VCG |
Morph |
|
|
Hôte |
CV. z |
||||||
|
G. |
GA (L) 13 |
Géorgie |
Feuille |
Gala |
G1 |
1 |
SP1 |
|
GA 16 |
Géorgie |
Fruit |
Gala |
G1 |
1 |
SP1 |
|
|
CROTTS (L) 27 |
Caroline du Nord |
Feuille |
Gala |
G1 |
1 |
SP1 |
|
|
TN 7 |
TN |
Feuille |
Gala |
G1 |
1 |
SP1 |
|
|
LD 11 |
Caroline du Nord |
Fruit |
GS |
G1 |
2 |
SP1 |
|
|
LD 16 |
Caroline du Nord |
Fruit |
GS |
G1 |
2 |
SP1 |
|
|
LD 25 |
Caroline du Nord |
Fruit |
GS |
G1 |
2 |
SP1 |
|
|
LD 20 |
Caroline du Nord |
Fruit |
GS |
G1 |
2 |
SP1 |
|
|
OH 3 |
OH |
Fruit |
MARYLAND |
G1 |
3 |
SP1 |
|
|
CROTTS 1 |
Caroline du Nord |
Fruit |
Gala |
G2 |
2 |
SP1 |
|
|
CROTTS 2 |
Caroline du Nord |
Fruit |
Gala |
G2 |
2 |
SP1 |
|
|
CROTTS 3 |
Caroline du Nord |
Fruit |
Gala |
G2 |
2 |
SP1 |
|
|
CROTTS 5 |
Caroline du Nord |
Fruit |
Gala |
G2 |
2 |
SP1 |
|
|
BR 2 |
Brésil |
Feuille |
Gala |
G3 |
5 |
SP1 |
|
|
BR 3 |
Brésil |
Feuille |
Gala |
G3 |
4 |
SP1 |
|
|
BR 8 |
Brésil |
Feuille |
Gala |
G3 |
4 |
SP1 |
|
|
BR 10 |
Brésil |
Feuille |
Gala |
G3 |
4 |
SP1 |
|
|
BR 13 |
Brésil |
Feuille |
Gala |
G3 |
5 |
SP1 |
|
|
BR 17 |
Brésil |
Feuille |
Gala |
G3 |
5 |
SP1 |
|
|
BR 19 |
Brésil |
Feuille |
Gala |
G3 |
5 |
SP1 |
|
|
BR 9 |
Brésil |
Feuille |
Gala |
G4 |
5 |
SP1 |
|
|
BR 21 |
Brésil |
Feuille |
Gala |
G4 |
5 |
SP1 |
|
|
RD 1 |
Caroline du Nord |
Fruit |
Délicieux |
A3 |
6 |
CP |
|
|
RD 2 |
Caroline du Nord |
Fruit |
Délicieux |
A3 |
6 |
CP |
|
|
LD 2 |
Caroline du Nord |
Fruit |
GS |
A3 |
6 |
CP |
|
|
LD 12 |
Caroline du Nord |
Fruit |
GS |
A3 |
6 |
CP |
|
|
TN 8 |
TN |
Feuille |
Gala |
A3 |
6 |
CP |
|
|
C. gloeo- |
AL 5 |
AL |
Fruit |
GD |
B2 |
7 |
SS2 |
|
AL 6 |
AL |
Fruit |
GD |
B2 |
7 |
SS2 |
|
|
AL 8 |
AL |
Fruit |
GD |
B2 |
7 |
SS2 |
|
|
AL 9 |
AL |
Fruit |
GD |
B2 |
7 |
SS2 |
|
|
AL 1 |
AL |
Fruit |
GD |
B3 |
8 |
SS3 |
|
|
AL 2 |
AL |
Fruit |
GD |
B3 |
8 |
SS3 |
|
|
AL 4 |
AL |
Fruit |
GD |
B3 |
8 |
SS3 |
|
|
AL 10 |
AL |
Fruit |
GD |
B3 |
8 |
SS3 |
|
|
LD Cg 1 |
Caroline du Nord |
Fruit |
GS |
B3 |
9 |
SS1 |
|
|
LD Cg 8 |
Caroline du Nord |
Fruit |
GS |
B3 |
9 |
SS1 |
|
|
LD Cg 11 |
Caroline du Nord |
Fruit |
GS |
B3 |
9 |
SS1 |
|
|
LD Cg 13 |
Caroline du Nord |
Fruit |
GS |
B3 |
9 |
SS1 |
|
|
C. |
LD Ca 5 |
Caroline du Nord |
Fruit |
GS |
C1 |
n / a |
SSC |
|
LD Ca 10 |
Caroline du Nord |
Fruit |
GS |
C1 |
n / a |
SSC |
|
|
LD Ca (b) 4 |
Caroline du Nord |
Fruit |
GS |
C1 |
n / a |
SSNC |
|
|
LD Ca (b) 6 |
Caroline du Nord |
Fruit |
GS |
C1 |
n / a |
SSNC |
|
|
BR Ca 4 |
Brésil |
Feuille |
Gala |
D1 |
13 |
SSNC (O) |
|
|
BR Ca 3 |
Brésil |
Feuille |
Gala |
D1 |
n / a |
SSNC (O) |
|
|
BR Ca 6 |
Brésil |
Feuille |
Gala |
D1 |
13 |
SSNC (O) |
|
x Des haplotypes d'ADN mt, des groupes de compatibilité végétative (VCG) et des types morphologiques ont été décrits dans une étude précédente (8).
y Abréviations: GA, NC, TN, OH et AL = Géorgie, Caroline du Nord, Tennessee, Ohio et Alabama, respectivement.
z GS = Granny Smith, MD = Molly's Delicious, GD = Golden Delicious.
Pour déterminer la température de croissance optimale, des bouchons de 5 mm de diamètre, situés à la périphérie de cultures âgées de 4 jours des isolats de G. cingulata, C. gloeosporioides et C. acutatum, ont été placés au centre de boîtes de Pétri contenant 15 ml de PDA et incubé dans le noir à 14, 18, 24, 26 et 30 ° C. Des bouchons de cinq mm de diamètre de chaque isolât ont été placés sur trois boîtes de PDA différentes et le diamètre des colonies a été mesuré après 2, 4 et 6 jours. L'expérience a été répétée une fois.
Analyses de données
La température optimale pour la croissance de chaque espèce a été estimée en ajustant une équation quadratique à la croissance de chaque espèce à toutes les températures testées [croissance = b0 + b1 (temp) + b2 (temp 2 )] et en résolvant l'équation suivante: croissance maximale = (-b1 / 2b2), où b1 et b2 sont les coefficients pour les termes linéaire et quadratique. La réduction de la croissance des isolats de G. cingulata, C. gloeosporioides et C. acutatum à des températures inférieures à l'optimum a été déterminée en calculant le pourcentage de réduction du diamètre de la colonie par rapport au diamètre de la colonie à la température où la croissance maximale s'est produite.
La croissance moyenne au sein des haplotypes et des espèces dans chaque expérience a été comparée à une analyse de variance utilisant SAS (version Windows, version 6.12; SAS Institute, Inc., Cary, Caroline du Nord). Les moyens ont été séparés par le test Waller-Duncan k -ratio t. La CE 50 (concentration efficace de bénomyl pour réduire la croissance de 50%) de chaque isolat en bénomyl a été calculée sur la base du diamètre moyen des colonies dans les haplotypes et les espèces 6 jours après l'incubation à l'aide du programme Proc Probit log10 de SAS.
Effet de la température et du bénomyl sur la croissance d'un isolat
Dans l’ensemble, les isolats de G. cingulata et de C. gloeosporioides ont augmenté plus rapidement et étaient plus sensibles au bénomyl que les isolats de C. acutatum (tableau 3). Il n'y avait pas de différences significatives dans le taux de croissance et la sensibilité au bénomyl entre les isolats de G. cingulataet C. gloeosporioides. Le taux de croissance entre les isolats de C. acutatumcollectés aux États-Unis (C1) et au Brésil (D1) était significativement différent, et les isolats de C. acutatum aux États-Unis ont augmenté plus lentement que tous les haplotypes testés.
Tableau 3. Taux de croissance et sensibilité au bénomyl des isolats de G. cingulata, C. gloeosporioides et C. acutatum
|
Espèce |
Nb de |
ADN mt |
Croissance (mm / jour) wy |
EC 50 xy |
||
|
Haplo- |
Espèce |
Haplo- |
Espèce |
|||
|
G. cingulata |
5 |
A3 |
13.0 a |
0,19 c |
||
|
G. cingulata |
7 |
G3 |
12.1 c |
0,12 dc |
||
|
G. cingulata |
4 |
G2 |
12,4 ba |
0,10 cc |
||
|
G. cingulata |
2 |
G4 |
12.3 av. |
- z |
||
|
G. cingulata |
9 |
G1 |
12.2 c |
12.4 a |
0,15 cc |
0,14 b |
|
C. gloeo- |
4 |
B2 |
12,9 ba |
0,16 cc |
||
|
C. gloeo- |
8 |
B3 |
11,4 d |
11.9 a |
0,07 j |
0,10 b |
|
C. acutatum |
4 |
C1 |
9.2 e |
0,37 b |
||
|
C. acutatum |
3 |
D1 |
8.5 f |
8.9 b |
0.66 a |
0.47 a |
v Les haplotypes d'ADN mt décrits dans une étude précédente (8).
w La croissance représente le diamètre de la colonie (mm / jour) des isolats à 26 ° C sur 6 jours.
x EC 50 s ont été calculés avec Proc Probit log 10 dans SAS, sur la base du diamètre de la colonie des isolats à 0, 0,01, 0,1, 1 et 10 mg / ml de bénomyl après 6 jours d'incubation à 26 ° C.
y Les moyennes suivies de la même lettre ne sont pas significativement différentes à P = 0,05 selon le test de Waller-Duncan k -ratio t.
z L' haplotype n'a pas été examiné.
C. acutatum avait une CE 50 plus élevée au bénomyl que les isolats de G. cingulata et C. gloeosporioides. Les isolats de C. acutatum du Brésil étaient les moins sensibles avec une CE 50 de 0,66 mg de bénomyl par ml (tableau 3).
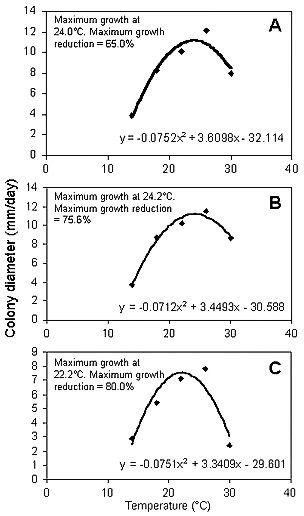
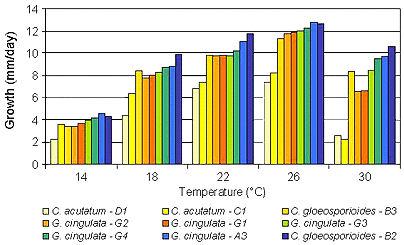
Les températures optimales prédites pour la croissance des isolats de G. cingulata, C. gloeosporioides et C. acutatum étaient respectivement de 24,0, 24,2 et 22,0 ° C; 14 ° C était la température la moins favorable pour toutes les espèces et la croissance à cette température a été réduite de 68 à 72% par rapport à la température optimale (Fig. 4). Les plus grandes différences de croissance entre les espèces ont été observées à 30 ° C (figure 5). À 30 ° C, les isolats de C. acutatum ont augmenté beaucoup plus lentement que ceux de G. cingulata et de C. gloeosporioides. A cette même température, des différences au sein d'isolats de la même espèce ont également été observées. Chez C. gloeosporioides B3, les isolats ont augmenté plus lentement que les isolats B2.Les isolats de G. cingulata présentant les haplotypes G1 et G2 ont augmenté plus lentement que ceux présentant les haplotypes G3, G4 et A3.
|
|
|
Fig. 4. Effet de la température sur la croissance des isolats de G. cingulata (A), C. gloeosporioides (B) et C. acutatum(C). Croissance maximale = (-b1 / 2b2), où b1 et b2 sont les coefficients des termes linéaire et quadratique, respectivement, dans l'équation quadratique [croissance = b0 + b1 (temp) + b2 (temp 2)]. La réduction de croissance maximale a été calculée à 14 ° C pour les trois espèces. |
|
Figure 5. Croissance des isolats de G. cingulata, C. gloeosporioides et C. acutatum à cinq températures différentes (14, 18, 22, 26 et 30 ° C). La croissance représente la moyenne sur 6 jours du diamètre moyen des colonies (mm / jour) des isolats mesuré tous les 2 à 6 jours. Les isolats testés représentent 9 haplotypes trouvés dans l'analyse RFLP de l'ADN mt et chaque haplotype est représenté par des isolats de différents endroits et des vergers de pommiers, le cas échéant (tableau 2). |
Conclusion
Dans des études antérieures, les isolats de G. cingulata et de C. gloeosporioides se distinguaient par leur taux de croissance plus rapide (10, 12, 14,18) et leur sensibilité accrue au bénomyl (1, 2, 12,18) à partir des isolats de C. acutatum obtenus de pommes, d'agrumes, de fraises, de pêches et d'autres hôtes. De plus, Adaskaveg et Hartin (1) ont observé une température de croissance optimale inférieure pour les isolats de C. acutatum obtenus à partir de fraises, d’amandes et de pêches, par rapport à la température de croissance optimale pour des isolats de C. gloeosporioides obtenus à partir d’agrumes et de papaye. Cependant, Gunnell et Gubler (10) ont déclaré que les isolats de G. cingulata, C. gloeosporioides et C. acutatum de fraise avaient la même température de croissance optimale. Dans cette étude, une croissance plus lente, une sensibilité moindre au bénomyl et une température de croissance optimale inférieure ont différencié les isolats de C. acutatum des isolats de G. cingulata et de C. gloeosporioides. Le taux de croissance et la sensibilité au bénomyl ne sont pas utiles pour distinguer les isolats de G. cingulata et de C. gloeosporioides, ni au sein des haplotypes d'ADN mt, des groupes de compatibilité végétative (GCV) ou des groupes morphologiques de chaque espèce. Des différences entre la croissance des haplotypes d'une même espèce n'ont été observées qu'à 30 ° C; cependant, seuls les isolats de G. cingulata qui causent la pourriture amère et GLS au sein de l'haplotype G1 ont été distingués des isolats de G. cingulata qui ne causent que la pourriture amère. Ces résultats suggèrent que, bien que les caractéristiques culturelles soient utiles pour différencier les espèces de Colletotrichum des pommes, elles ne sont pas utiles pour séparer les isolats en dessous du niveau de l'espèce.
Date de dernière mise à jour : 11/04/2023
Ajouter un commentaire