
Tylenchulus semipenetrans
Tylenchulus semipenetrans (nématode des agrumes, nématode des racines des agrumes ou anguillule du citronnier) est une espèce cosmopolite de nématodes de la famille des Tylenchulidae, originaire d'Extrême-Orient.
Ce nématode est un ravageur qui cause des dégâts significatifs dans toutes les régions agrumicoles du monde. Il attaque également d'autres plantes cultivées comme la vigne, l'olivier, le kaki, le lilas...
Tylenchulus semipenetrans
Tylenchulus semipenetrans
(Cobb, 1913)
HISTORIQUE. RÉPARTITION GÉOGRAPHIQUE. . ORIGINE
Un dépérissement des Citrus observé en Californie, fut pour la première fois attribué à un nématode par THOMAS (1913). L'agent, Tylenchulus semipenetrans, est décrit par COBB la même année (1913). Mais ce parasite existait déjà dans d'autres contrées puisque très rapidement il est signalé par COBB (1914) sur des Citrus en provenance de Floride, Espagne, Malte, etc. ., et par TRABUT (1915) en Algérie. En fait, T. semipenetrans est pratiquement présent partout où des plantations étendues d'agrumes existent. Il a été signalé à ce jour dans les pays suivants : Europe : Espagne, France, Italie, Malte, U. R. S. S. Afrique : Afrique du Sud, Algérie, Égypte, Maroc, Rhodésie, Uganda, Tunisie. Amérique : Argentine, Brésil, Chili, États-Unis, Pérou. (*) Ingénieur agronome, Institut F. de Rech. Fruitières O. M ., Centre de Côte d'Ivoire, Abidjan . (**) Maître de Recherches O. R. S. T. O. M. Institut d'Enseignement et de Recherches Tropicales, Abidjan, Côte d’Ivoire. - 4 45 Asie : Ceylan, Chine, Formose, Israël. Océanie : Australie, Hawaii. T. semipenetrans vient d'être récemment reconnu par les auteurs en Côte-d'Ivoire dans de vieilles plantations de pamplemoussiers et d'orangers ainsi qu'en Guinée. Il est certain que ce parasite existe dans bien d'autres contrées. Étant donné la vaste répartition géographique de ce parasite due vraisemblablement aux transports de plants de Citrus effectués de tous temps, il peut sembler vain de rechercher son lieu d'origine. Toutefois CHITWOOD et BIRCHFIELD (1957) supposent que T. semipenetrans est originaire de Floride ; ces deux auteurs l'ont en effet rencontré en plusieurs occasions sur Mihania batati f olia D. C ., une Composée sauvage très répandue dans cette région ; plusieurs des prélèvements positifs provenaient de lieux qui n'avaient jamais été mis en culture.
LE PARASITE
Les neufs de T. semipenetrans éclosent dans le sol. Les larves qui en sortent ont déjà subi une mue à l'intérieur de l'œuf comme il est de règle chez le groupe (Tylenchida) auquel appartient cette espèce. Ces larves sont donc dites « larves de 2e stade ». Elles se D FIG. r. - Stades libres de Tylenchulus serrai,-penetrans Cobb. A : Larve femelle, 2e stade. - B : Larve mâle, 2e stade. - C : Mâle adulte. D : Jeune femelle adulte. - (d'après S. D. Van Gundy, 1958). LE PARASITE Fruits - Vol. 16, no 9, 1961 présentent sous la forme d'un fuseau, arrondi à l'avant et assez effilé vers l'arrière, long de 300 µ à 350 ; elles possèdent un stylet long de 12 à 13 t orné de trois boutons basaux arrondis ; l'œsophage comprend un bulbe médian valvulaire moyennement développé suivi d'une assez longue partie mince (l'isthme) se terminant par un bulbe basal glandulaire renflé ne recouvrant pas l'intestin . C'est ce dernier caractère qui permet d'identifier aisément les larves de T. semipenetrans lors des comptages routiniers et de les différencier notamment des larves de Meloidogyne souvent présentes dans les mêmes échantillons : alors que chez ces dernières la limite œsophage-intestinale est très oblique et peu nette, chez les larves de T. sein-penetrans cette limite, très distincte, est perpendiculaire au grand axe du corps. En cas de doute, un examen microscopique à fort grossissement révélera le caractère typique du genre : la position du pore excréteur situé sur la moitié postérieure du corps alors que chez tous les autres genres de Tylenchida il est situé au plus au niveau du quart antérieur. Il n'y a pas à craindre de confusion avec d'autres espèces du même genre ; la seule autre espèce connue, Tylenchulus mangenotii, strictement localisée et spécialisée, parasite en effet une plante sauvage en de rares points de la Côte d'Ivoire. Dès ce stade, on peut reconnaître les larves qui donneront des femelles de celles qui donneront des mâles les premières sont plus minces et plus effilées à l'arrière que les secondes et ont un œsophage plus long. L'évolution des larves mâles et des larves femelles est très différente (VAN GUNDY, 1958). Les larves mâles demeurent très peu de temps au second stade (2 à 3 jours) et subissent rapidement trois autres mues pour se transformer en mâle adulte. L'ensemble du processus depuis l'éclosion de l'œuf peut ne prendre que 18o heures. Les mâles adultes, longs de 330 µ. à 410 µ, possèdent un œsophage réduit non fonctionnel ; le stylet n'est pratiquement plus visible ; les organes sexuels par contre sont très développés et le testicule unique remplit plus du tiers du corps. Ne pouvant se nourrir et vivant sur leurs réserves les mâles doivent avoir une durée de vie courte. Pendant toute la durée de leur développement ils vivent dans le sol ; le développement entier peut se passer en l'absence d'hôte ; les mâles ne sont donc pas des parasites obligés. Les larves femelles peuvent demeurer par contre très longtemps au deuxième stade qui constitue le stade de résistance du nématode ; cette durée dépend essentiellement de la température du sol et peut varier de deux ans et demi pour un sol à 15° C à deux mois et demi pour un sol à 32,5° C (BAINES, 1956).
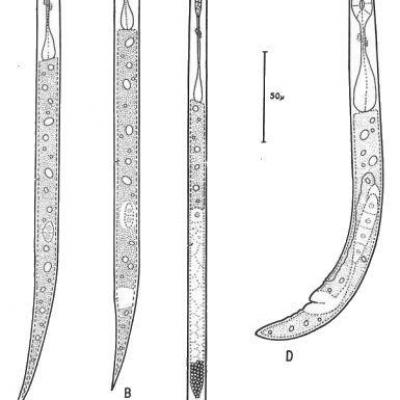
FIG. 1. - Stades libres de Tylenchulus semipenetrans Cobb. A : Larve femelle, 2e stade. - B : Larve mâle, 2e stade. - C : Mâle adulte. D : Jeune femelle adulte.
Avant de se transformer en femelles adultes les larves de deuxième stade subissent elles aussi trois mues successives. Au cours de ces mues leur morphologie change la queue se raccourcit et s'arrondit, le corps s'épaissit, l'œsophage se développe en longueur et en largeur ; les jeunes femelles sont plus trapues avec une queue courte, la portion postérieure du corps arquée ventralement, un œsophage atteignant presque la moitié du corps ; les lèvres de la vulve, située très postérieurement, se renflent. Entre la deuxième mue et la quatrième conduisant à l'adulte, il s'écoule environ sept jours. Les larves femelles ne semblent pas pénétrer dans les racines, mais vivent à la surface de celles-ci ; les jeunes femelles pénètrent ensuite assez rapidement dans les racines. La partie antérieure du corps, jusqu'après la jonction œsophage-intestinale, s'insère entre les cellules du parenchyme radiculaire, la partie postérieure restant à l'extérieur de la racine, d'où le nom de l’espèce. Cette partie postérieure se gonfle considérablement par suite du grand développement pris par la gonade femelle. A pleine maturité la femelle se présente avec une partie antérieure d'abord effilée, droite, puis montrant des renflements très variables en nombre et en forme, car ils proviennent du moulage de l'animal dans les espaces intercellulaires. Cette portion antérieure contient le stylet, l'œsophage, pourvu d'un volumineux bulbe médian 'à valvules très développées, faisant office de pompe aspirante et foulante, et un bulbe basal glandulaire à trois noyaux. La portion extérieure du corps est en forme de cornemuse, renflée, avec une queue courte, arquée ventralement. L'appareil génital comprend une seule gonade, très contournée. Il n'y a pas d'anus visible, par contre l'appareil excréteur (en compensation ?) est bien développé et le pore excréteur situé près de la vulve. La cuticule de la partie extérieure est nettement plus épaisse que celle de la partie engagée dans les tissus radiculaires. Une seule femelle peut pondre un assez grand nombre d'oeufs (75 à 100). Ces oeufs mêlés à une substance gélatineuse restent agglomérés en masse -autour de la femelle en formant de petites boules irrégulières, auxquelles adhèrent des particules de sol. A l'intérieur de ces masses se trouvent non seulement des oeufs, mais aussi des larves, des mâles et de jeunes femelles, car le développement peut commencer immédiatement après la ponte. Le cycle d'œuf à veuf a une durée de six à huit semaines. La fécondation des femelles a lieu alors que celles-ci n'ont pas encore pénétré dans les racines. Cette fécondation n'est d'ailleurs pas obligatoire, et des femelles non fécondées peuvent pondre des oeufs contenant des larves, mâles et femelles, parfaitement viables. L'infestation des Citrus par T. semipenetrans est sous la dépendance étroite de la température du sol (BAINES, 1950) : la plage optimum s'étend de 25° C à 31° C ; à 15° et à 35° l'infestation ne sera que très légère. Quand la température du sol descend en hiver au-dessous de 15°, les larves entrent en état de quiescence, demeurent inactives mais ne meurent pas. Tous les stades de T. semipenetrans sont très sensibles à la sécheresse ; par contre, les larves et les mâles survivent très bien à une submersion prolongée pendant plus de neuf mois (GUTIERREZ, 1947). De même l'action de différents sels solubles (NaCl, SO4NA2, CaCl2) conduit à un accroissement de la population de T. senti-penetrans (MACHMER, 1958).
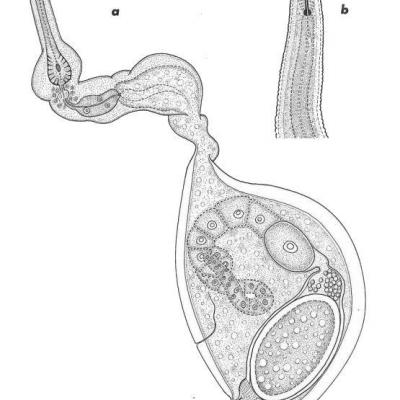
Tylenchulus semipenetrans
a) Femelle adulte mâture
b) Détail de la tête.
Classification :
Date de dernière mise à jour : 30/10/2023
Ajouter un commentaire