
LA TIGE
Sur un arbre, nous ne rencontrons aucune difficulté pour reconnaître une tige principale ou tronc, des tiges secondaires, branches et rameaux mis en place au cours des années successives. De même, le stipe (du latin stipes = tige) des Fougères arborescentes, des Cycadales et des Palmiers, non ramifié, sensiblement cylindrique et sur lequel les bases de feuilles tombées restent bien visibles, se distingue fort bien d’un tronc, abondamment ramifié à partir d’une certaine hauteur au-dessus du sol, de forme tronconique et ne présentant pas de restes de bases foliaires.
En fait, la tige est un organe très polymorphe, comme le montreront les exemples cités plus loin. Aussi devons-nous rechercher une définition pouvant concerner tous les types caulinaires (du grec caulos = tige) des Spermaphytes et être étendue aux Ptéridophytes, comme aux Bryophytes.
1/ La ramification
De l’importance et de la distribution des différents éléments de l’ensemble caulinaire dépend le port d’un individu, c’est-à-dire son aspect général, sa silhouette. Ce port est en relation avec l’importance de la ramification d’une part, l’intensité de la croissance en longueur et en épaisseur d’autre part. Comme celle des Thallophytes, la ramification des Cormophytes est dichotomique ou latérale.
- La dichotomie vraie (du grec dikha = en deux et tomê = section)
Très répandue chez les Ptéridophytes, elle se réalise au moment où le sommet de la tige se scinde en deux moitiés, chacune commandant alors la croissance en longueur des deux branches de la fourche qui vient de se former. Malgré ce partage, chacun des deux sommets-fils récupère rapidement l’importance du sommet d’origine, si bien qu’il n’existe pas de variations sensibles de diamètre d’un étage à l’autre d’un système dichotomique vrai.
Ramification dichotomique, isotone (en haut) et anisotone (en bas)
La taille des rameaux-fils est égale (ramification isotone – du préfixe iso et du grec tonos = tension), ou inégale (ramification anisotone – du grec anisos = inégal).
Les dichotomies se font successivement dans le même plan (des Sélaginelles et des Lycopodes) ou se croisent alternativement à angle droit (dichotomie rectangulaire des Lépidodentracées, Lycopodinées arborescente des marécages anthracolithiques de l’Hémisphère Nord, de Sélaginelles, de Lycopodes et de Fougères fossiles.)
- La ramification latérale
Caractéristique des Spermaphytes, mais se rencontrant aussi chez les Ptéridophytes (Articulées) et les Bryophytes (mousse, Hépatiques à feuilles), elle a son siège, non seulement à l’extrémité distale de la tige comme dns le cas précédent, mais aussi latéralement et à différents niveaux. D’une tige principale se détachent alors des rameaux latéraux.
- Place des bourgeons
- Chez les Spermaphytes, c’est à partir d’un bourgeon axillaire situé sur la tige et à l’aisselle d’une feuille (position adaxiale) que se forme un rameaux. En revanche, chez des Ptéridophytes (Articulées), les rameaux latéraux apparaissent entre deux feuilles. Quant à ceux des Mousses, ils naissent en position dorsale (ou inférieure) par rapport à la feuille et non en position ventrale (ou supérieure) comme dans le cas des Spermaphytes. Cette ramification latérale des Articulées et des mousses est en rapport avec la position particulière des bourgeons, entre les feuilles et adossés à l’axe dans le premier cas, en position abaxiale dans le second.
- Les bourgeons adventifs apparus (Un organe végétatif est qualifié d’adventif quand, s’ajoutant secondairement à d’autres organes du même type, il est d’une origine et occupe une position différente) par exemple, sur la section d’un tronc d’arbre après cicatrisation et à partir desquels se forment des rejets, n’ont aucune position particulière vis-à-vis de l’insertion des anciennes feuilles tombées depuis plusieurs années. De même, d’autres bourgeons adventifs portés par des racines produisent des drageons, tiges sortant du sol à une certaine distance de l’individu qui les a engendrées (arbres fruitiers, herbes vivaces).
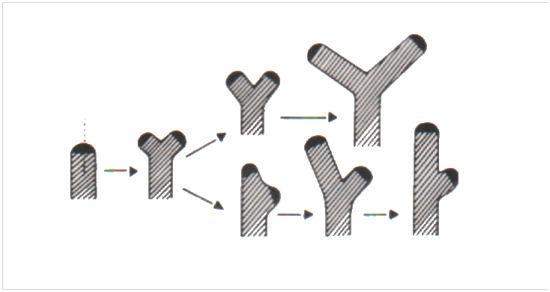
- Monopole et sympode

Ramification sympodiale monochasiale : A, à l’automne, la partie distale du rameau de l’année, son bourgeon terminal et un bourgeon latéral dégénèrent ; B, un bourgeon latéral s’est redressé et est maintenant en position terminale. Il est flanqué de deux cicatrices : à gauche, celle résultant de la chute du sommet desséché du rameau et, à droite, celle de la feuille tombée à l’aisselle de laquelle ce bourgeon se trouvait à l’origine en position latérale (les flèches indiquent les cicatrices foliaires).
- La ramification des Spermaphytes présente deux variantes. Elle est dite monopodiale quand le bourgeon terminal de la tige principale (et celui des ramifications nées de bourgeon axillaires) reste fonctionnel d’une année sur l’autre. La tige principale et ses ramifications sont donc constituées d’unités annuelles résultant de l’activité périodique d’un seul et vrai bourgeon terminal. Si la ramification est sympodiale, le bourgeon se trouvant apparemment en position terminale sur la tige principale et ses ramifications est, en réalité, un bourgeon axillaire redressé. En effet chaque unité annuelle est produite par un bourgeon latéral venu se placer secondairement en position terminale après la chute du sommet de l’unité précédente. Tous ces bourgeons ne fonctionnent donc qu’une fois. A propos de la ramification sympodiale, on dit qu’elle est monochasiale quand un seul bourgeon axillaire redressé est en cause, et dichasiale si deux bourgeons insérés en vis-à-vis sont concernés. Bien que, dans ce dernier cas, une fourche se forme, cette ramification latérale particulière ne peut être confondue avec la ramification dichotomique. Ces deux types de ramification latérale ne s’excluent pas, une espèce pouvant être monopodiale avant floraison et sympodiale après. C’est le cas du Marronnier (Hippocastanacées).
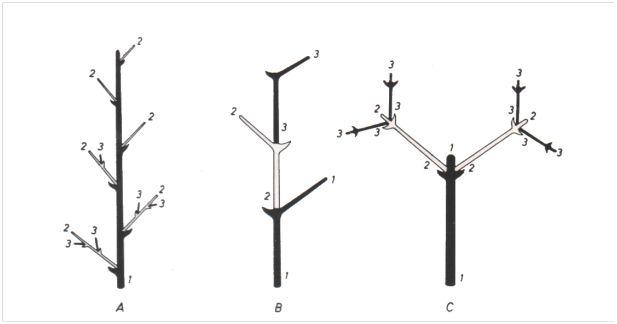
Ramification latérale : A, monopodiale ; B, sympodiale monochasiale ; C, sympodiale dichasiale. Les chiffres indiquent l’ordre d’apparition des axes.
- Si, à première vue, ramification dichotomique et ramification latérale paraissent bien différentes, les ramifications monopodiales pourraient pourtant dériver, comme le suggère Emberger, de la ramification dichotomique. En effet, quand celle-ci est anisotone, la feuille angulaire est déjetée, soit du côté défavorisé, soit du côté favorisé. Le résultat rappelle alors une ramification monopodiale dans le premier cas, sympodiale dans le second. Divers Ptéridophytes illustrent ces deux possibilités, si bien que la ramification latérale axillaire vraie n’est connue, dans ce groupe, que chez une Filicinée fossile du genre Ankyropteris (du Carbonifère moyen au Permien inférieur).
- Monocaulie et multicaulie
L’appareil végétatif des espèces monocaules est constitué d’une tige unique, axe non ramifié, dressé, dont le développement dépend du fonctionnement d’un seul méristème apical (stipe en colonne des grands Palmiers). L’appareil végétatif des multicaules (= polycaules), au contraire, est constitué d’un système d’axes résultant de l’activité coordonnée de bourgeons végétatifs axillaires.

Rapports entre la ramification dichotomique anisotone et la ramification latérale : à gauche, rameau de Selaginella apus avec feuilles angulaires (fa) en position de bissectrice (pr : porte-racines = rhizophore) : à droite, effets théoriquement possible du déplacement de la feuille angulaire d’une ramification dichotomique (a) pouvant expliquer les rapports entre la dichotomie et les ramifications monopodiale monochasiale (c).
L’organisme monocaule, à la merci d’un traumatisme de son seul centre de croissance, ne survit pas à la destruction de son méristème apical. Au contraire, chez les espèces multicaules, le grand nombre de foyers de ramification (à l’extrémité et le long de l’axe central et des axes latéraux) fait que toute mutilation est compensé par l’activité de bourgeons de remplacement. L’architecture monocaule est donc beaucoup moins compétitive vis-à-vis de la sélection naturelle que la multicaule.
Monocaulie et multicaulie coexistent depuis les temps les plus reculés puisque les Psilophytinées fossiles étaient déjà ramifiées. D’ailleurs, tout multicaule est obligatoirement monocaule au début de sa vie quand il est à l’état de plantule. La ramification est un phénomène secondaire qui, suivant les espèces, s’installe précocement ou tardivement.
2/ La croissance
D’une manière générale, la longueur des tiges augmente plus rapidement que leur diamètre au cours de la croissance.
- La croissance en longueur
L’allongement total, très variable d’une espèce à l’autre (pins : 30 cm/an, Bambou : 60 cm/jour) est le résultat de croissances différentielles, car toutes les parties d’une tige ne s’allongent pas également. Les nœuds, au niveau desquels sont insérées les feuilles, s’allongent peu, alors que les entre-nœuds (les supérieurs en particulier) subissent, au contraire, une élongation appréciable. C’est pourquoi cette croissance est qualifiée d’intercalaire.
Bien que la croissance en longueur des tiges soit théoriquement indéfinie pour des raisons physiologiques encore mal élucidées la taille des Cormophytes est limitée. (Un organe est dit à croissance indéfinie quand il garde la possibilité de croître pendant toute sa vie, aux seules périodes d’activité bien entendu, cette croissance n’étant pas continue. Celle-ci est au contraire définie, quand elle ne se fait sentir qu’au début de l’existence de l’organe. Elle s’interrompt définitivement assez tôt, si bien que celui-ci acquiert rapidement la taille qu’il conservera pendant toute son existence. C’est le cas de la feuille).
La croissance en longueur de la tige s’accompagne d’un mouvement révolutif, dit de circumnutation, de son extrémité (on désigne par mutation tout mouvement de croissance non orienté par rapport à un facteur excitant.), mouvement d’une grande amplitude chez les plantes volubiles, ce qui leur permet de rencontrer un support autour duquel elles s’enroulent.
L’orientation des tiges en croissance est également sous la dépendance de l’attraction terrestre. Aussi, oppose-t-on souvent le géotropisme négatif de la tige au géotropisme positif de la racine. En fait, le comportement de ces organes est beaucoup plus complexe que semble l’indiquer l’image d’une tige dressée vers le ciel et d’une racine dirigée en sens inverse. En effet, s’il existe des tiges orthotropes (du grec orthos = droit) effectivement verticales (orthogéotropisme), plus ou moins indifférents à la pesanteur, sont plagiotropes (du grec plagios = oblique), obliques ou horizontales (diagéotropisme, du grec dia = à travers, et gê = terre). Au cours de la vie de la plante, ces deux géotropismes ne sont pas forcément constants pour un même rameau. Ainsi, au moment de sa mise en place, une pousse de Pin de l’année est douée d’un orthogéotropisme négatif net, puis elle s’infléchit pour s’aligner avec la branche oblique qui l’a formée.
Les tiges orthotropes et les tiges plagiotropes se rencontrent chez les Bryophytes et les Ptéridophytes, comme chez les Spermaphytes.
- La croissance en épaisseur
Dans la flore actuelle, seuls des Spermaphytes (Gymnospermes et Angiospermes Dicotylédones) présentent une croissance en épaisseur importante, souvent même rapide, plus spécialement les plantes arborescentes que les plantes herbacées. En revanche, parmi les Ptéridophytes fossiles, les Lycopodinées avec les Lépidodendracées, et les Articulées avec les Calamitacées, ont montré une croissance en épaisseur souvent spectaculaire, leur ayant permis de réaliser des arbres atteignant de 1 à 2 mètres de diamètre.
Le rythme de l’accroissement circulaire est le même que celui de la croissance intercalaire, l’intensité de ces deux phénomènes étant maximum, en général, en juin et en juillet sous nos latitudes.
3/ Les principaux ports
- Le port arborescent
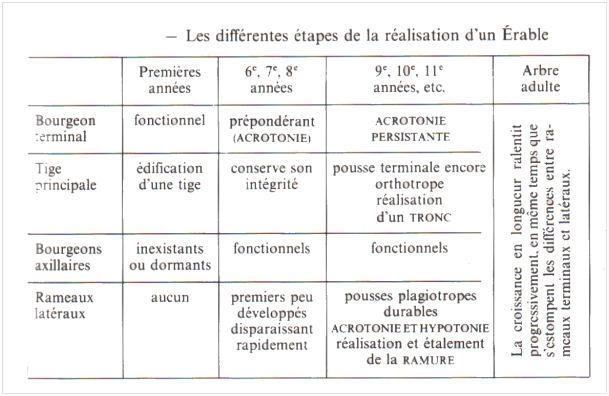
L’acrotonie (du grec akros = extrême et tonos = tension) est caractérisée par la prépondérance du bourgeon terminal sur tous les bourgeons axillaires et celle des bourgeons axillaires les plus proches du sommet de la tige sur ceux qui en sont éloignés. D’où l’établissement d’un gradient morphologique basipète régulièrement décroissant se matérialisant par le volume de plus en plus petit des bourgeons le long de la tige principale (ou des rameaux latéraux), et par la taille de plus en plus courte des pousses qu’ils engendrent.
Si l’acrotonie se manifeste sur la tige principale comme sur les ramifications plagiotropes, l’hypotonie (du préfixe hypo = en dessous) est particulièrement à ces dernières. Les bourgeons de la face inférieure des divers rameaux dominent ceux de la face supérieure ; il en est de même des pousses correspondantes, d’où l’étalement progressif de la ramure.
Dans le cas particulier des Érables, chaque nœud porte toujours deux feuilles diamétralement opposées, une paire foliaire faisant un angle de 90° avec les deux paires immédiatement voisines (feuilles opposées décussées). Il en résulte que l’hypotonie ne se manifeste pleinement avec anisophyllie et anisocladie (inégalité des feuilles et des rameaux insérés sur un même nœud, du grec phullon = feuille et kladôdês = rameau), que tous les deux nœuds, quand l’une des feuilles (et le rameau correspondant) est en position supérieure, l’autre étant obligatoirement en position inférieure. Au contraire, tous les deux nœuds également, il y aura isophyllie et isocladie (du grec isos = égal).
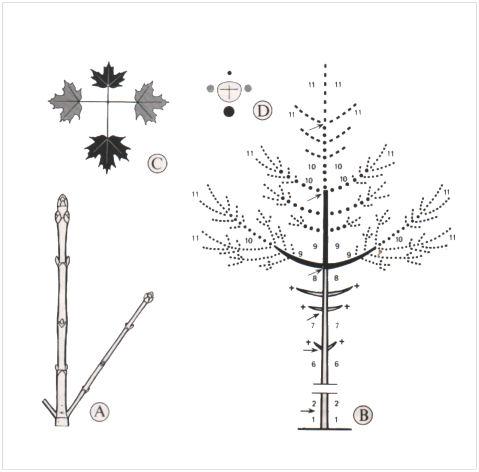
Le port arborescent du Sycomore : A, la taille respective des bourgeons met en évidence l’acrotonie et hypotonie ; B, agencement des ramifications successives mises en place pendant les onze premières années (les croix indiquent que les rameaux latéraux des 6e, 7e et 8e années disparaissent au moins partiellement. Les flèches limitent les unités annuelles de la tige principale et les nombres de 1 à 11 se rapportent aux unités mises en place chaque année sur la tige principale, comme sur les ramifications, d’un Sycomore de onze ans) ; C, anisophyllie entre deux couples de feuilles opposées décussées d’un rameau latéral de l’année ; D, coupe transversale schématique de ce rameau montrant sa dorsiventralité hypotone accompagnée d’anisocladie suivant les nœuds considérés. Dans la silhouette générale du schéma B, les petits rameaux relevant de l’isocladie n’ont pas été figurés.
Arbres et arbustes sont, en principe, les uns monopodiaux (Érables et Pinacées), les autres sympodiaux (Châtaigniers, Ormes, Tilleuls), mais les notions de monopode et de sympode ne représentent pas des différences capitales, car ces deux types de ramification n’influent pas sur le port arborescent commandé essentiellement par l’acrotonie d’abord et secondairement par l’hypotonie. En effet, celle-ci est quelquefois remplacée par une amphitonie (du grec amphi = autour), les bourgeons les plus forts étant situés sur les flancs des rameaux latéraux. L’étalement de la ramure persiste, mais toutes les ramifications latérales d’une même branche sont placées sensiblement dans un même plan. Il en résulte que la silhouette d’un Épicéa (Gymnospermes/Pinacées), par exemple, est d’apparence beaucoup plus régulière que celle d’un Érable.
- Le port buissonnant
La figure ci-dessous montre comment se réalise progressivement un buisson d’Épine-vinette (Berberis vulgaris/Berbéridacées), de Groseillier (Ribes/Saxifragacées) ou de divers Rosiers (Rosa/Rosacées).

Le port buissonnant : agencement des ramifications successives mises en place pendant les quatre premières années. Une coupe transversale des rameaux montre que leur dorsiventralité est épitone.
Alors que la partie inférieure d’un tronc d’arbre est dépourvue de branches (ceci en rapport avec l’acrotonie), la basitonie entraîne, au contraire, l’apparition de nombreuses ramifications à la base d’un buisson. D’autre part, si l’hypotonie était responsable de l’étalement de la ramure par la mise en place de rameaux latéraux plagiotropes, l’épitonie se traduit, au contraire, par la formation de rameaux orthotropes au cœur du buisson, qui devient ainsi de plus en plus fourni au cours des années. Ce double renversement de dominance oppose donc, en principe, le type arbre au type buisson.
- Le port herbacé
Ce port est plus varié que les précédents, l’inhibition des bourgeons axillaires étant totale (d’où absence de ramifications latérales), ou seulement partielle. Dans ce cas, les rameaux les plus longs sont insérés au sommet ou à la base de la tige principale, suivant les espèces.
Sur ces thèmes existent de nombreux cas particuliers parmi lesquels :
- La ramification rythmique des Caryophyllacées chez qui, à chaque nœud, un des deux bourgeons évolue en pousse longue, l’autre restant totalement inhibé ou ne fournissant qu’une pousse de petite taille.
- Chez les graminées, les bourgeons bien développés au niveau des nœuds de la base du chaume produisent des pousses secondaires, c’est le tallage. Il en résulte la mise en place d’une touffe serrée au niveau du sol (port graminiforme en rapport avec la ramification basilaire). Si l’on couche de jeunes chaumes, on accentue le tallage naturel en provoquant la formation de pousses secondaires au niveau d’entre-nœuds qui n’en produiraient pas normalement. C’est pourquoi on préconise, par exemple, de rouler le gazon des pelouses plusieurs fois par an afin d’accentuer l’implantation des touffes des différentes espèces qui le constituent.
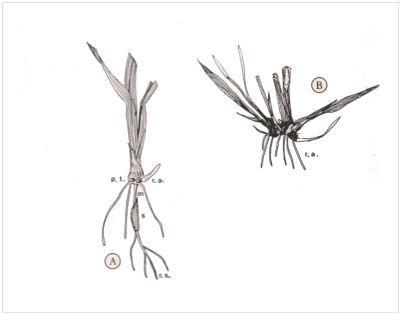
Tallage naturel de l’Arrhenatherum elatius (graminées) : A, plantule (s, semence ; m, mésocotyle ; r.s., racines séminales ; plateau de tallage ; r.a., racines adventives) ; B, plusieurs talles se sont formés au niveau du plateau tandis que semence, mésocotyle et racines séminales ont disparu. Dans le cas présent, chez la sous-espèce bulbosum, le nœud basal de chaque talle se tubérise.
- Complexité des problèmes physiologiques posés par le port d’une plante
L’architecture des Cormophytes est sous la dépendance de corrélations diverses entre le bourgeon apical et les bourgeons axillaires, entre les cotylédons ou les feuilles et leurs bourgeons axillaires respectifs. Il en résulte des inhibitions ou des stimulations de croissance variables d’une espèce à l’autre et, pour un même individu, avec son âge, l’époque de l’année, l’intensité lumineuse, l’importance de la compétition. Aussi avons-nous dit précédemment que l’acrotonie et l’hypotonie d’une part, la basitonie et l’épitonie d’autre part, opposent, en principe seulement, le port arborescent au port buissonnant. C’est ainsi que chez le Sureau noir (Sambucus nigra/Caprifoliacées), la pousse feuillée herbacée produite au printemps présente une nette acrotonie. Sa croissance cesse à la fin de l’été et elle devient basitone. La basitonie d’un rameau d’un an s’est substituée à une acrotonie préexistante. La période de repos apparent d’un végétal est donc importante pour l’établissement de nouveaux rapports entre les bourgeons.
Conséquence directe de telles observation : le notion de port, bien qu’étant descriptive, n’en est pas moins dynamiques, car elle peut rendre compte des transformations qui, au court de la vie d’un végétal, en modifient la silhouette.
4/ La diversité des types caulinaires
Pour un même port, deux espèces ou un individu peuvent, suivant les cas, posséder des rameaux de taille et de forme très différentes en fonction de l’importance relative de leurs croissances en longueur et en épaisseur.
Nous signalerons ici les types caulinaires spécialisés les plus répandus concernant la tige principale, comme ses rameaux latéraux, en empruntant plus particulièrement nos exemples aux Spermaphytes, chez qui le polymorphisme caulinaire est le plus riche.
- Les plantes acaules
A premières vue, il semble qu’une Joubarbe (Crassulacées) ne possède pas de tige. En fait, une coupe longitudinale met en évidence une très courte tige qui s’allonge lors de le floraison. D’une manière générale, les plantes dites en rosettes, aux feuilles plus ou moins plaquées sur le sol, comportent une tige de très petite taille qui peut échapper à l’observation, au moins au cours de la phase végétative et même, pour certaines espèces, pendant toute la vie de l’individu. Ce sont des plantes acaules.

Joubarbe avant floraison : à gauche, seules les feuilles charnues et la racine sont visibles ; à droite, une coupe longitudinale de l’individu fait apparaître la tige aplatie.
- Les tiges grêles
- Les lianes ne restent dressées que si elles s’accrochent à un support (espèces grimpantes comme la Vigne et le Lierre) ou s’enroulent autour (espèces volubiles comme le Houblon et la Clématite). Les Préphanérogames actuels et les Gymnospermes ne sont jamais lianoïdes. Très rare chez les Ptéridophytes (Equisetum giganteum d’Amérique est une liane de 12 mètres pour un diamètre de 0,5 à 2 cm), les lianes sont surtout répandues parmi les Angiospermes.
- Les stolons sont des tiges horizontales, aux longs entre-nœuds, aux feuilles très réduites et différentes de celles portées par la tige principale, courte et verticale (les fraisiers).

Stolon de Fraisiers : en s’enracinant, le stolon émis par la plante-mère a produit un individu-fils.
- Les chaumes sont également des tiges grêles, mais relativement moins longues et plus rigides que les lianes et les stolons. Ces tiges noueuses (les nœuds sont très saillants), généralement creuses au niveau des entre-nœuds, caractérisent des Graminées.
- Les tiges de petite taille
- Cas particulier des bourgeons
Ce sont des rameaux dont la croissance, temporairement interrompue, sera maximum au moment de leur débourrement. Un bourgeon est une tige encore plus courte que celle d’une rosette et dont les ébauches de nœuds et de feuilles sont, le plus souvent, recouvertes d’écailles.
Dans le cas des bourgeons typiquement axillaires, il arrive :
- Que la plupart des aisselles foliaires soient vides (Gymnospermes) ;
- Que des bourgeons soient cachés par la base dilatée du pétiole (Platanes) ;
- Qu’il existe des bourgeons accessoires surnuméraires alignés verticalement.
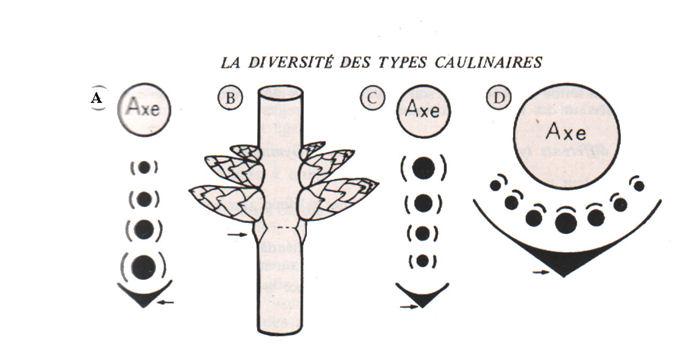
Les bourgeons accessoires surnuméraires : A et B, bourgeons sériaux ascendants ; C, bourgeons sériaux descendants ; D, bourgeons collatéraux (les flèches indiquent les feuilles ou leur emplacement). Pour chaque bourgeon, on observe deux préfeuilles (ou premières feuilles réduites à l’état d’écailles, le plus souvent) situées de part et d’autre du plan médian de la feuille (Dicotylédones), ou une préfeuille unique adossée à l’axe (Monocotylédones).
(Bourgeons sériaux descendants ou ascendants des Dicotylédones), ou situés dans un plan horizontal au niveau d’un nœud (bourgeons collatéraux des Monocotylédones). Le cas du Robinier (Robinia pseudacacia/Légumineuses) mérite une mention particulière car chez cet arbre les bourgeons sont à la fois sériaux et cachés. En effet au nombre de deux à cinq pour chaque feuille, enfermés dans une cavité axillaire remplie de nombreux poils roux, ils ne sont pas vivibles à l’extérieur avant le printemps suivant. Ils sortent alors par une fente de la cavité et le bourgeon le plus élevé s’épanouit. Si la pousse formée vient à geler, un bourgeon inférieur prend le relais et débourre à son tour.

Les bourgeons sériaux du Robinier. Le scalp de la tige a eu pour conséquence le débourrement de deux bourgeons cachés qui ont mis en place deux rameaux latéraux (1 et 2) (s : stipule)
- Que la forme des bourgeons soit variable chez un même individu, suivant qu’ils sont situés sur des rameaux ordinaires ou sur des rejets (Peupliers).
- Les différents types rameaux des Pinacées (Gymnospermes)
Suivant leur taille, on distingue :
- Des rameaux longs (ou auxiblastes) à croissance indéfinie, avec bourgeon terminal et bourgeons axillaires fonctionnels ;
- Des rameaux courts (ou mésoblastes) à croissance limitée dans le temps et dans l’espace, avec bourgeon terminal et quelques bourgeons axillaires rarement fonctionnels.
- Des rameaux nains (ou brachyblastes) à croissance très limitée, avec bourgeon terminal, mais sans bourgeons latéraux.
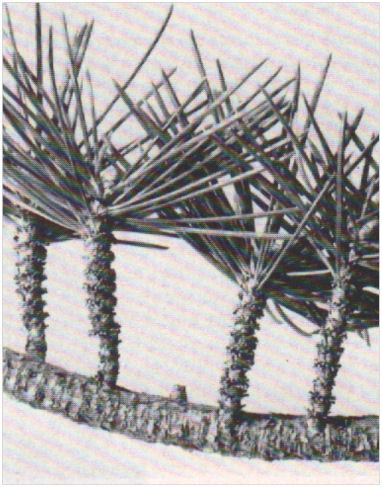
Les rameaux courts de cèdre. Disposées sur des rameaux longs, chacun d’eux porte à son sommet un bouquet de feuilles aciculaires rigides.
Le tableau ci-dessous indique, pour des Conifères (Pins, Cèdres et Épicéas), la répartition de ces trois types de rameaux et de leurs feuilles (écailles ou aiguilles), les genres Pinus, Cedrus et Picea marquant les étapes d’une série phylogénique progressivement réductionnelle.

Les rameaux courts ne sont pas l’apanage des Gymnospermes. Le Hêtre (Fagus silvatica/Fagacées), ainsi que des arbres fruitiers de la famille des Rosacées, en possèdent également.
- Rameaux spécialisés à croissance définie
- Les rameaux-épines ou dards. Apparus au printemps, leur croissance en longueur s’arrête bientôt et leur bourgeon terminal durcit. La pointe acérée de l’épine est réalisée, tandis que les feuilles sous-jacentes se fanent et tombent. Seules les feuilles basales persistent plus longtemps si le dard est assez long, leurs bourgeons axillaires donnant des rameaux-épines secondaires (Aubépine-Crataegus, Argousier-Hippophae rhamnoides, Ajoncs-Ulex.). Les rameaux-épines ne doivent pas être confondus avec les aiguillons de certaines tiges (ronces, rosiers), simples hypertrophies localisées des tissus périphériques caulinaires.
- Les rameaux-vrilles. Les lianes grimpantes des Vitacées, en particulier celles de la Vigne, se fixent à un support grâce à des vrilles de nature caulinaire, rameaux courts volubiles.
- Les cladodes. Ce sont des rameaux courts constitués d’un seul entre-nœud, mais toujours d’une grande ressemblance avec des feuilles, qu’ils soient aplatis (cladodes de Ruscus aculeatus), ou en aiguilles (cladodes des Asparagus). Malgré cette convergence morphologique flagrante, les cladodes sont axillées par des feuilles (réduites et écailleuses), et peuvent porter des bourgeons et des fleurs comme les rameaux ordinaires.
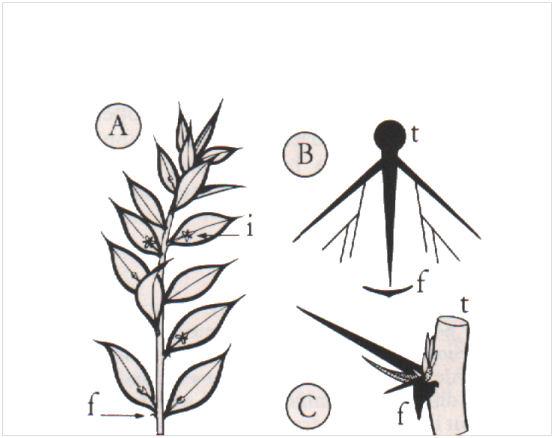
Les cladodes : A, un rameau long de Ruscus aculeatus portant, à l’aisselle de feuilles écailleuses (f), des cladodes aplatis sur lesquels se développent des inflorescences (i) ; B (diagramme et C (vue de profil) d’un fascicule de cladodes aciculaires ‘Asparagus (ramification d’abord dichasiale, puis monochasiale) (t : tige ; f : feuille écailleuse).
- Des rhizomes aux bulbes
Constituant tout ou partie du système caulinaire de certains Cormophytes, ces organes, le plus souvent souterrains et gorgés de réserves, contribuent à la conservation des espèces. D’une résistance particulière, ils supportent la mauvaise saison en vie ralentie, alors que les parties aériennes ont pu disparaître totalement. C’est sous forme de rhizomes, de tubercules ou de bulbes que les géophytes invisibles sous nos latitudes pendant la période hivernale, n’en restent pas moins en vie. En se ramifiant, puis en se fragmentant, ces tiges particulières assurent aussi une multiplication végétative active.
- Les rhizomes
Tiges plus ou moins plagiotropes dans la majorité des cas, portant des feuilles écailleuses et des racines adventives hypotones, les rhizomes produisent à la belle saison des rameaux orthotropes aériens feuillés et florifères ou seulement des feuilles également aériennes.
Suivant le nombre et la taille des entre-nœuds qui constituent les rhizomes, on distingue des rhizomes :
- Longs (plusieurs mètres), assez minces, à croissance rapide : beaucoup de plantes de nos dunes littorales comme l’Oyat (Ammophila arenaria/Graminées) et le Carex arenaria (Cypéracées). Quand, dans les dunes fixées, des surfaces ont été mises à nu par le vent, le Carex arenaria « recoud » la dune en lançant, sur de grandes distances, les différentes ramifications de son rhizome. Bien que celles-ci ne soient pas visibles, on peut les suivre littéralement à la trace, du fait de la présence de chapelets d’axes aériens feuillés qui les jalonnent sur toute leur longueur. Si l’on met à jour l’une de ces ramifications, on constate qu’elle peut atteindre plusieurs mètres (quelquefois plus de dix), ceci en rapport avec une croissance en longueur active, et qu’elle porte un très grand nombre d’axes aériens. Dans les dunes vives, l’Oyat a un comportement comparable, ce qui lui vaut d’être utilisé dans la fixation du sable.
- Moins longs, d’un diamètre supérieur, à croissance moins rapide : Muguet (Convallaria maialis/Liliacées).
- Plus ou moins hypertrophiés, à entre-nœuds courts, à propagation relativement lente : Polygonatum multiflorum (Liliacées), beaucoup d’Iris (Iridacées).

Rhizome de Polygonatum multiflorum.
Chaque année le rhizome met en place une unité nouvelle qui porte une tige aérienne florifère. Celle de l’année en cours de l’échantillon photographié a été sectionnée, tandis que celles des deux années précédentes sont disparues, mais la zone circulaire de leur insertion est encore bien visible.
Variable d’une espèce à l’autre, la profondeur à laquelle croissent les rhizomes est relativement constante pour une espèce donnée. Cette propriété explique, en partie, la répartition des espèces dunaires rhizomateuses du littoral en fonction de l’importance de l’ensablement local.
- Les tubercules
Renflement localisés de la tige principale ou de ses ramifications, généralement souterrains, les tubercules caulinaires (Le terme de tubercule est également employé pour des racines hypertrophiées et des organes mixtes) sont formés par des stolons ou des rhizomes. Leurs feuilles sont écailleuses.
Les réserves des tubercules sont fréquemment utilisées par l’Homme pour son alimentation. Dans la famille intertropicale des Dioscoréacées, les tubercules souterrains, ou même aériens, sont très répandus et souvent exploités pour leurs réserves amylacées.
L’Igname (Dioscorea batatas), par exemple, est cultivée comme légume en Asie.
Nous citerons parmi :
- Les tubercules de stolons, ceux de la pomme de terre (Solanum tuberosum/Solanacées). Chacun des yeux visibles à leur surface est une dépression, allongée transversalement, dont le bord inférieur plus abrupt que le supérieur, porte une feuille écailleuse très réduite et rapidement caduque. Le fond de cette dépression est occupé par le bourgeon axillaire de cette feuille, souvent flanqué de deux bourgeons secondaires plus petits. Chez le rhizome de Topinambour (Helianthus tuberosus/Composées), les feuilles sont persistantes.
- Les tubercules de rhizomes, ceux du Crosne du Japon (Stachys sieboldii/Labiacées) et du Cyperus esculentus (Cypéracées).
- Les tubercules hypocotylaires provenant de l’hypertrophie de l’hypocotyle, partie de la tige de l’embryon située sous les cotylédons, ceux du Radis (Raphanus sativus/Crucifères).
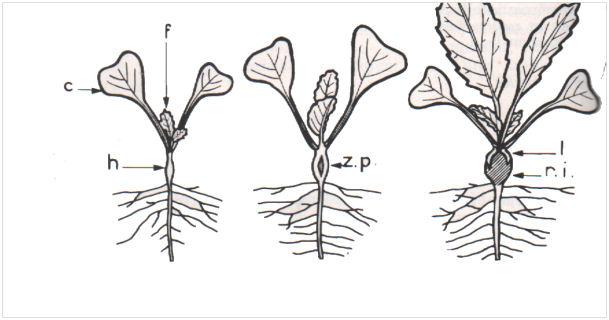
Trois étapes de la tubérisation de l’hypocotyle de radis (c, cotylédons ; f, feuille ; h, hypocotyle ; z.p, zone périphérique de l’hypocotyle déchirée par la tubérisation de la région interne (r.i) ; l : languettes subsistant quelque temps après la déchirure.
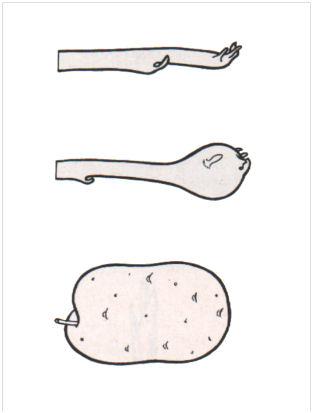
Trois étapes de la tubérisation de l’extrémité d’un stolon de pomme de terre.
- Les bulbes
Ces tiges le plus souvent orthotropes, larges et plus courtes que les rhizomes, portent des feuilles en général bien développées et dont les parties souterraines ne sont pas chlorophylliennes.
Suivant l’importance relative de la tige et des feuilles, on distingue des bulbes caulinaires et des bulbes foliaires.
Constitués, dans le premier cas, d’une tige hypertrophiée enveloppée seulement par des bases foliaires minces et membraneuses, ils sont denses et qualifiés de solides.
La famille des Iridacées montre bien le passage du rhizome typique des Iris constitué de plusieurs unités plagiotropes d’années différentes, au bulbe rhizomateux du Glaïeul encore plagiotrope, mais réduit à l’unité de l’année et au bourgeon de l’année suivante, puis au bulbe des Crocus encore plus court et devenu orthotrope.

Bulbe rhizomateux plagiotrope de Glaïeul (A), bulbe solide orthotrope de Crocus vernus (B) et de Crocus sativus en groupe longitudinale (C) (b1 : bulbe de l’an dernier ou des années précédentes ; b² : bulbe de l’année ; r : racines adventives ordinaires ; r.t. : racines adventives tractives ; f.j. : jeunes feuilles ; f.m. : bases membraneuses d’anciennes feuilles ; b.a. : bourgeon apical ; a.t. : axe tubérisé ; r.a. : racines adventives.
- Les tiges succulentes
Dans la famille des cactacées, les tiges sont presque toujours succulentes, c'est-à-dire gorgées d’eau (du grec succos = plein de suc). Quelle que soit leur forme, elles développent une surface relativement faible par rapport à leur volume. Des convergences morphologiques sont frappantes entre les tiges des Cereus (Cactacées), de l’Euphorbia echinus (Euphorbiacées), des Stapelia (Asclépiadacées) et du Senecio anteuphorbium (Composées). Aussi parle-t-on d’une manière générale de tiges cactiformes et de plantes cactoïdes.
Date de dernière mise à jour : 18/04/2022
Ajouter un commentaire