
MONOCOTYLÉDONE.
Monocotylédone
Lilianae, Liliopsida, Monocotyledonae
Les Monocotylédones, Monocotyledonae, classe des Liliopsida selon la classification de Cronquist ou le super-ordre des Lilianae selon les classifications APG III et APG IV, sont un groupe monophylétique qui comprend, parmi les angiospermes ou plantes à fleurs, des végétaux dont la plantule typique présente un seul cotylédon sur l'embryon évoluant en une préfeuille (ou éophylle), contrairement aux Dicotylédones qui ont deux.
On relève notamment parmi les monocotylédones :
- les Orchidées, qui présentent des fleurs parmi les plus spécialisées pour la pollinisation,
- les Palmiers ou Arecaceae,
- les Bananiers (genre Musa),
- les Graminées ou Poacées,
- les Joncs, de la famille des Juncaceae.
Lilianae

{kind=link}
Jeune pousse d’ Iris pseudacorus
|
Classification selon Tropicos |
|
Caractéristiques morphologiques
À leurs deux synapomorphies principales (un seul cotylédon et plastes des tubes criblés du type P II c) s'ajoutent les caractéristiques suivantes du faciès monocotyloïdes :
- racines : appareil racinaire souvent fasciculé, c'est-à-dire constitué de racines adventives d'égale importance et généralement non ramifiées. L'endoderme des racines de monocotylédones est en "fer à cheval", c'est-à-dire que les cellules de cette assise sont lignifiées sur les parois anticlines et sur la paroi péricline interne, donnant une forme de U en coupe transversale. La voie apoplastique est ainsi bloquée. Certaines cellules sont tout de même non lignifiées et permettent la circulation de l'eau et des solutés vers le cylindre central, elles sont appelées cellules de passage.
- tiges : pas de formation de bois secondaire et absence d'un véritable tronc ; même si certaines monocotylédones (palmiers, bananiers, Pandanus, Cordylines, Yuccas, Dracénas, Bambous, ravenala, phynakospermum, dasylirion,…) ont un port arborescent, on ne rencontre pas dans cette classe de vrais arbres au sens strict (tous sont des feuillus et des conifères, qui sont tous des dicotylédones et des gymnospermes), la tige des palmiers étant appelée un stipe. Les faisceaux libéro-ligneux sont implantés sur plusieurs cercles concentriques, à l'inverse des dicotylédones, pour lesquels la disposition de ces faisceaux est uniquement sur un seul cercle. Des faisceaux de fibres faites de cellules de sclérenchyme allongées donnent une grande résistance à la traction.
- feuilles : habituellement simples et allongées, parfois pennées, rarement composées (Arisaema), présentant le plus souvent (la parisette, la salsepareille, le tamier, la Goodyère rampante font l'exception) des nervures parallèles, leurs connexions latérales plutôt peu nombreuses étant peu ramifiées ; ces feuilles ont constitué, selon des botanistes, des phyllodes (pétioles aplatis en limbe selon la théorie du phyllode pétiolaire de de Candolle en 1827, supportée par Agnes Arber en 1918 mais désormais réfutée)9.
- fleurs : fondamentalement trimères : 3 sépales, 3 pétales, 2 × 3 étamines, 3 carpelles ; symétrie dont on retrouve la trace même chez des fleurs évoluées comme celles des orchidacées.
- pollen : grains de pollen possédant généralement une seule aperture, zone de faiblesse permettant le passage du tube pollinique, caractère plésiomorphique partagé avec les dicotylédones ancestrales.
Les Monocotylédones sont essentiellement herbacées, moins de 10 % des espèces ont une tige plus ou moins ligneuse avec une sorte de cambium vascularisé typique des plantes ligneuses non-dicotylédones. Les plantes monocotylédones ligneuses, contrairement aux dicotylédones ligneuses, ont généralement une tige non ramifiée avec une couronne terminale de grandes feuilles. Les scientifiques supposent fortement que les caractéristiques morphologiques sont liées à large adoption d'un habitat aquatique par les différentes lignées de plantes monocotylédones. Les éléments vasculaires manquent souvent ou sont parfois limités aux organes spécifiques. Les faisceaux vasculaires de la tige sont généralement dispersés ou supportés en deux cycles ou plus. Le système racinaire mature est adventif. Un seul cotylédon, ou feuille embryonnaire indifférenciée, est présent. Les plantes monocotylédones possèdent généralement des feuilles avec des nervures parallèles. Les pièces florales sont essentiellement de type ternaire (fleurs trimères), rarement quaternaire (fleurs tétramères) et quasi jamais quinaire (fleurs pentamères). Le pollen est essentiellement monoaperturé, une seule aperture par grain.
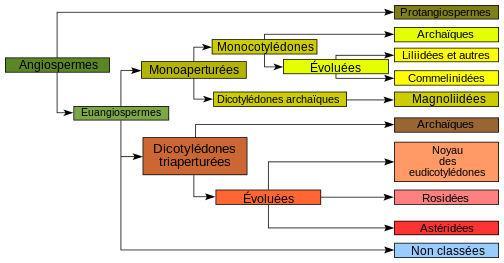
Classifications
{kind=link}
Les Monocotylédones sont apparues il y a 130 millions d’années et ont un ancêtre commun avec les eudicotylédones. D’un point de vue évolutif, les travaux récents (2009) 10,11 en biologie moléculaire appliquée à la cladistique permettent de positionner les monocotylédones entre les angiospermes basales (par exemple les Nymphaeales) et les eudicotylédones. Lorsque les botanistes distinguent Monocotylédones et Dicotylédones, ils caractérisent davantage des types d'organisation que des unités systématiques naturelles, sachant que les classifications verticales (par lignées) sont d'un tout autre intérêt que les classifications horizontales (par convergence). Bien que la très grande majorité des plantes actuelles soient des dicotylédones, les monocotylédones sont pourtant les végétaux les plus évolués : le plus ancien angiosperme, qui est une dicotylédone, a pour plus proche parent actuel l'amborella.
Classification classique de Cronquist
En classification classique de Cronquist (1981), les monocotylédones sont la classe Liliopsida dans les divisions Magnoliophyta [= angiospermes]. Elles sont divisées en 5 sous-classes :
- classe des Liliopsida
- sous-classe des Alismatidae
- sous-classe des Arecidae
- sous-classe des Commelinidae
- sous-classe des Liliidae
- sous-classe des Zingiberidae
Classification classique de Thorne
En classification classique de Thorne (1992), les monocotylédones sont la sous-classe des Liliidae dans la classe des Magnoliopsida [= angiospermes]. Elles sont divisées en 8 superordres.
Classifications APG
En classification phylogénétique APG III (2009), les monocotylédones (« monocots » en anglais) forment un clade situé dans le clade angiosperme (« angiosperms » en anglais). Elles sont divisées en 11 ordres (et une famille). Leur composition et leur emplacement sur l'arbre phylogénétique du vivant sont détaillés par le cladogramme suivant :
- groupe des Mésangiospermes (Mesangiospermae, en anglais « mesangiosperms »)
-
- Chloranthales
- clade des Magnoliidées (Magnoliidae, en anglais « magnoliids »)
-
- clade des Monocotylédones (en anglais « monocotyledons » ou « monocots »)
- Acorales
-
- Alismatales
-
- Petrosaviales
-
-
- Liliales
-
- Asparagales
- clade des Commelinidées (Commelinidae, en anglais « commelinids »)
- clade des Monocotylédones (en anglais « monocotyledons » ou « monocots »)
-
- Dasypogonaceae
- Arecales
- Poales
-
- Zingiberales
- Commelinales
-
- Ceratophyllales
- clade des Eudicotylédones ou Dicotylédones vraies (en anglais « eudicots » ou « eudicotyledons »)
-
Remarque :
- Dans la première classification phylogénétique APG (1998), le nom anglais commelinoids (Commélinoïdes) était utilisé pour les commelinids (Commélinidées) de la classification phylogénétique APG III (2009).
- L'ordre des Petrosaviales est un ajout d'APG III contenant la famille Petrosaviaceae auparavant placée sans ordre.
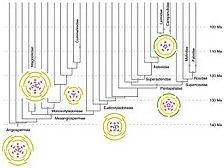
Évolution simplificatrice
{kind=link}
Évolution florale depuis 140 Ma, représentée par les diagrammes floraux.
Dès la fin du XIXe siècle, des botanistes comme George Henslow (en) considèrent que les Monocotylédones, en raison principalement de leur anatomie plus simple, sont des dégradations des Dicotylédones, cette évolution étant due à l'habitat aquatique de leurs ancêtres. La communauté des botanistes a depuis réfuté cette origine dicotylédonique mais l'évolution simplificatrice due à l'habitat aquatique des ancêtres des Monocotylédones fait consensus : ces ancêtres, peut-être des géophytes hygrophiles ou amphibies auraient émergé dans des forêts humides à canopée fermée. Les structures rigides, en raison de la poussée d'Archimède, ne sont plus nécessaires et seuls des organismes de tailles réduites, comme des herbes, peuvent se développer dans les lacs et rivières ; l'absorption de l'eau par l'ensemble des parties immergées cantonne les racines à un rôle d'ancrage ; les feuilles deviennent rubanées ; la disparition des structures secondaires (cambium, phellogène) entraîne l'absence de ramification (à l'exception des inflorescences légères, cette absence expliquant la perte du second cotylédon), l'absence de port arborescent, de racine pivotante (développement de racines adventives), de feuille composée, de liège (d'où le développement de géophytes pour passer la mauvaise saison).
Liste des ordres
Selon ITIS (11 octobre 2017) :
- ordre des Acorales
- ordre des Alismatales
- ordre des Arecales
- ordre des Asparagales
- ordre des Commelinales
- ordre des Dasypogonales
- ordre des Dioscoreales
- ordre des Liliales
- ordre des Pandanales
- ordre des Petrosaviales
- ordre des Poales
- ordre des Zingiberales
Selon Tropicos (11 octobre 2017)
- ordre des Acorales Mart.
- ordre des Agrostidales Tagliana, Giulio
- ordre des Alismatales R. Br. ex Bercht. & J. Presl
- ordre des Arecales Bromhead
- ordre des Asparagales Link
- ordre des Commelinales Mirb. ex Bercht. & J. Presl
- ordre des Cyclanthales Poit.
- ordre des Cyperales Hutch.
- ordre des Dasypogonales Doweld
- ordre des Dioscoreales Mart.
- ordre des Flagellariales Takht. ex Reveal & Doweld
- ordre des Hypoxidales Takht. ex Reveal & Doweld
- ordre des Juncales Dumort.
- ordre des Liliales Perleb
- ordre des Lowiales Takht. ex Reveal & Doweld
- ordre des Monocotylondoneae E. Morren ex Mez
- ordre des Musales Reveal
- ordre des Najadales Dumort.
- ordre des Orchidales Bromhead
- ordre des Pandanales R. Br. ex Bercht. & J. Presl
- ordre des Petrosaviales Takht.
- ordre des Poales Small
- ordre des Tecophilaeales Traub ex Reveal
- ordre des Triuridales Hook. f.
- ordre des Xanthorrhoeales Takht. ex Reveal & Doweld
- ordre des Xyridales Lindl. ex Arn.
- ordre des Zingiberales Griseb.
Selon World Register of Marine Species (11 octobre 2017) :
- ordre des Acorales
- ordre des Alismatales
- ordre des Arecales
- ordre des Asparagales
- ordre des Commelinales
- ordre des Dasypogonales
- ordre des Dioscoreales
- ordre des Liliales
- ordre des Pandanales
- ordre des Petrosaviales
- ordre des Poales
- ordre des Zingiberales
Date de dernière mise à jour : 20/01/2024
Ajouter un commentaire