
Puccinia graminis - Rouille de la tige
Les rouilles de la tige , du noir et des céréales sont causées par le champignon Puccinia graminis et sont une maladie importante affectant les cultures céréalières . Les espèces de cultures touchées par la maladie comprennent le blé tendre , le blé dur , l' orge et le triticale . Ces maladies ont affecté la culture des céréales à travers l'histoire. La récurrence annuelle de la rouille noire du blé dans les plaines du nord de l'Inde a été découverte par le professeur KC Mehta. Depuis les années 1950, des souches de blé sélectionnées pour résister à la rouille des tiges sont devenues disponibles. [3] Fongicidesefficace contre la rouille de la tige sont également disponibles.
|
Rouille de la tige |
|
|
Royaume: |
|
|
Division: |
|
|
Classe: |
|
|
Ordre: |
|
|
Famille: |
|
|
Genre: |
|
|
Espèce: |
P. graminis |
|
Puccinia graminis Pers. , (1794) |
|
|
Dicaeoma anthistiriae |
|

En 1999, une nouvelle race virulente de rouille noire a été identifiée contre laquelle la plupart des souches de blé actuelles ne montrent aucune résistance. La race a été nommée TTKSK (par exemple isolat Ug99 ), du nom du pays où elle a été identifiée ( Ouganda ) et de l'année de sa découverte (1999). Il s'est propagé au Kenya , puis en Éthiopie , au Soudan et au Yémen et devient de plus en plus virulent à mesure qu'il se propage. Une épidémie de rouille noire du blé causée par la race TTKSK se propage actuellement en Afrique , en Asie et au Moyen-Orient et est une source de préoccupation majeure en raison du grand nombre de personnes qui dépendent du blé pour leur subsistance. Les scientifiques travaillent sur la sélection de souches de blé résistantes à l'UG99. Cependant, le blé est cultivé dans un large éventail d'environnements. Cela signifie que les programmes de sélection auraient encore beaucoup de travail à faire pour obtenir une résistance dans des germoplasmes adaptés au niveau régional, même après l'identification de la résistance. De même, en 2014, la race apparentée à Ug99 appelée «Digalu», a également émergé et dévasté la variété Digalu en Éthiopie.
Une épidémie d'une autre race virulente de rouille noire, TTTTF, a eu lieu en Sicile en 2016, suggérant que la maladie est de retour en Europe. Une analyse génomique complète de Puccinia graminis combinée à des données sur la pathologie végétale et le climat a mis en évidence le potentiel de réémergence de la rouille du blé-tige au Royaume-Uni.
Contenu
Taxonomie

Modèle d'une spore de puccinia graminis , fin du xixe siècle, Musée botanique de Greifswald
Il existe une diversité génétique considérable au sein de l'espèce P. graminis et plusieurs formes spéciales, forma specialis , dont la gamme d'hôtes varie, ont été identifiées.
- Puccinia graminis f. sp. avenae , avoine
- Puccinia graminis f. sp. dactylis
- Puccinia graminis f. sp. hordei , orge
- Puccinia graminis f. sp. lolii
- Puccinia graminis f. sp. poae
- Puccinia graminis f. sp. secalis , seigle , orge
- Puccinia graminis f. sp. tritici , blé , orge
P. graminis est membre du phylum Basidiomycota au sein du royaume Fungi . La couleur rouille caractéristique des tiges et des feuilles est typique de la rouille générale de la tige ainsi que de toute variation de ce type de champignon. Différentes de la plupart des champignons, les variations de la rouille ont cinq stades de spores et alternent entre deux hôtes. Le blé est l'hôte principal et l' épine - vinette est l'hôte alternatif.
Il existe plusieurs pathotypes (y compris le QCC et le MCC) affectant l' orge , au sein de forma specialis tritici .
Pathologie
Le champignon de la rouille de la tige attaque les parties de la plante qui sont au-dessus du sol. Les spores qui se posent sur les plants de blé vert forment une pustule qui envahit les couches externes de la tige. Les plantes infectées produisent moins de talles et produisent moins de graines, et en cas d'infection grave, la plante peut mourir. L'infection peut réduire ce qui est une culture apparemment saine environ trois semaines avant la récolte en un enchevêtrement noir de tiges cassées et de grains ratatinés à la récolte.
La rouille de la tige des céréales entraîne des pertes de rendement de plusieurs manières:
- Le champignon absorbe les nutriments qui seraient autrement utilisés pour le développement du grain.
- Les pustules traversent l'épiderme, ce qui perturbe le contrôle de la transpiration de la plante et peut entraîner une dessiccation et une infection par d'autres champignons.
- L'interférence avec le tissu vasculaire végétal conduit à des grains ratatinés.
- Le champignon affaiblit les tiges, ce qui peut entraîner la verse (chute). Dans les cas graves, la verse peut rendre la récolte mécanique impossible.
Signes et symptômes
Sur le blé

Feuilles infectées par le blé avec un pathogène de la rouille des tiges avec un gène de résistance spécifique
La rouille de la tige sur le blé est caractérisée par la présence d'urédinia sur la plante, qui sont des pustules rouge brique, allongées, en forme de cloques qui sont facilement secouées. Ils se produisent le plus souvent sur les gaines foliaires, mais se trouvent également sur les tiges , les feuilles , les glumes et les arêtes . Sur les feuilles, ils se développent principalement sur la face inférieure mais peuvent pénétrer à la face supérieure. Sur les gaines foliaires et les glumes, des pustules rompent l'épiderme, donnant un aspect irrégulier. Vers la fin de la saison de croissance, des télias noires sont produites. Pour cette raison, la rouille de la tige est également connue sous le nom de «rouille noire». Les telia sont fermement attachés au tissu végétal.
Le site de l'infection est un symptôme visible de la maladie.
Sur l'épine-vinette
Les pycnia apparaissent sur les plants d'épine-vinette au printemps, généralement sur la face supérieure des feuilles. Ils sont souvent en petits groupes et exsudent des pycniospores dans un miellat collant. Cinq à dix jours plus tard, des structures en forme de coupe remplies d' éciospores poudreuses jaune orangé traversent la surface inférieure des feuilles. Les cupules éciales sont jaunes et parfois allongées pour s'étendre jusqu'à 5 mm de la surface des feuilles.
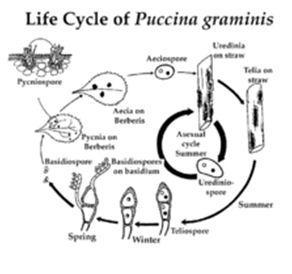
Cycle de la vie
{kind=link}
_-Osterloh-_-Brendel_10_h,_2-.jpg&usg=ALkJrhjjMK_uvZgYeVimvBZCaVqA1k48vQ){kind=link}
{kind=link}
{kind=link}
Cycle de vie de Puccinia graminis
Comme les autres espèces de Puccinia , P. graminis est un biotrophe obligatoire (il colonise les cellules végétales vivantes) et a un cycle de vie complexe caractérisé par une alternance de générations . Le champignon est hétéroïque , ce qui nécessite deux hôtes pour compléter son cycle de vie - l'hôte céréalier et l'hôte alternatif. Il existe de nombreuses espèces dans Berberis et Mahonia (et leur genre hybride x Mahoberberis ) qui sont sensibles à la rouille de la tige, mais l' épine - vinette commune est considérée comme l'hôte alternatif le plus important. P. graminis est macrocyclique (présente les cinq types de spores connus pour les champignons de la rouille).
Puccinia graminis peut compléter son cycle de vie avec ou sans épine-vinette (l'hôte alternatif).
Cycle de vie sur l'épine-vinette
En raison de sa nature cyclique, il n'y a pas de véritable «point de départ» pour ce processus. Ici, la production d’urédospores est choisie arbitrairement comme point de départ.
Les Urediniospores se forment dans des structures appelées urédinia, qui sont produites par des mycéliums fongiques sur l'hôte céréalier 1 à 2 semaines après l'infection. Les urédospores sont dicaryotes (contiennent deux noyaux haploïdes non fusionnés dans une cellule) et sont formées sur des tiges individuelles dans l' uredinium. Ils sont épineux et rouge brique. Les Urediniospores sont le seul type de spores dans le cycle de vie du champignon de la rouille qui sont capables d'infecter l'hôte sur lequel elles sont produites, et cela est donc appelé le «stade répétitif» du cycle de vie. C'est la propagation des urédospores qui permet à l'infection de se propager d'une plante céréalière à une autre. Cette phase peut rapidement propager l'infection sur une vaste zone.
Vers la fin de la saison de croissance de l'hôte céréalier, les mycéliums produisent des structures appelées telia. Telia produit un type de spore appelé téliospores. Ces spores noires à parois épaisses sont dicaryotes . Ils sont la seule forme sous laquelle Puccinia graminis est capable d'hiverner indépendamment d'un hôte. [11]
Chaque téliospore subit une caryogamie (fusion de noyaux) et une méiose pour former quatre spores haploïdes appelées basidiospores . C'est une source importante de recombinaison génétique dans le cycle de vie. Les basidiospores sont à parois minces et incolores. Ils ne peuvent pas infecter l'hôte céréalier, mais peuvent infecter l'hôte alternatif (généralement l'épine-vinette). Ils sont généralement transportés vers l'hôte alternatif par le vent.
Une fois que les basidiospores arrivent sur une feuille de l'hôte alternatif, elles germent pour produire un mycélium haploïde qui pénètre directement dans l'épiderme et colonise la feuille. Une fois à l'intérieur de la feuille, le mycélium produit des structures d'infection spécialisées appelées pycnia. La pycnie produit deux types de gamètes haploïdes, les pycniospores et les hyphes réceptifs. Les pycniospores sont produites dans un miellat collant qui attire les insectes. Les insectes transportent des pycniospores d'une feuille à l'autre. Les gouttes de pluie éclaboussantes peuvent également propager des pycniospores. Une pycniospore peut féconder un hyphe réceptif du type d'accouplement opposé, conduisant à la production d'un mycélium dicaryote . C'est le stade sexuel du cycle de vie et la fertilisation croisée fournit une source importante de recombinaison génétique.
Ce mycélium dicaryote forme alors des structures appelées écies , qui produisent un type de spores dicaryotes appelées éciospores . Celles-ci ont une apparence moelleuse et sont formées en chaînes - contrairement aux urédospores qui sont épineuses et sont produites sur des tiges individuelles. Les chaînes des éciospores sont entourées d'une enceinte en forme de cloche de cellules fongiques. Les éciospores sont capables de germer sur l'hôte céréalier mais pas sur l'hôte alternatif (elles sont produites sur l'hôte alternatif, qui est généralement l'épine-vinette). Ils sont transportés par le vent vers l'hôte céréalier où ils germent et les tubes germinatifs pénètrent dans la plante. Le champignon pousse à l'intérieur de la plante sous forme de mycélium dicaryote. En 1 à 2 semaines, le mycélium produit de l'urédine et le cycle est terminé.
Cycle de vie sans épine-vinette
Puisque les urédospores sont produites sur l'hôte céréalier et peuvent infecter l'hôte céréalier, il est possible que l'infection passe d'une récolte d'une année à l'autre sans infecter l'hôte alternatif (épine-vinette). Par exemple, les plants de blé spontanés infectés peuvent servir de pont d'une saison de croissance à une autre. Dans d'autres cas, le champignon passe entre le blé d'hiver et le blé de printemps, ce qui signifie qu'il a un hôte céréalier toute l'année. Puisque les urédospores sont dispersées par le vent, cela peut se produire sur de grandes distances. Notez que ce cycle consiste simplement en une multiplication végétative - les urédospores infectent un plant de blé, conduisant à la production de plus d'urédospores qui infectent ensuite d'autres plants de blé.
Dispersion des spores
Puccinia graminis produit les cinq types de spores connus pour les champignons de la rouille .
Les spores sont généralement déposées près de la source, mais la dispersion sur de longues distances est également bien documentée. Les trois catégories suivantes de dispersion à longue distance sont connues:
- Dispersion sur de très longues distances
- Cela peut se produire sans aide (la nature robuste des spores leur permet d'être transportées sur de longues distances dans l'air, puis déposées par lavage par la pluie) ou assistée (généralement sur des vêtements humains ou du matériel végétal infecté qui est transporté entre les régions). Ce type de dispersion est rare et est très difficile à prévoir.
- Extension de gamme par étapes
Il s'agit probablement du mode de dispersion à longue distance le plus courant et se produit généralement dans un pays ou une région.
- Extinction et recolonisation
Cela se produit dans les zones qui ont des conditions inadaptées à la survie toute l'année de Puccinia graminis - généralement des régions tempérées où les hôtes sont absents en hiver ou en été. Les spores hivernent ou passent l'été dans une autre région, puis se recolonisent lorsque les conditions sont favorables.
Gènes de résistance à la rouille de la tige du blé
Un certain nombre de gènes de résistance à la rouille des tiges (gènes Sr) ont été identifiés dans le blé. Certains d'entre eux sont nés dans le blé panifiable (par exemple Sr5 et Sr6 ), tandis que d'autres ont été élevés à partir d'autres espèces de blé (par exemple Sr21 de T. monococcum ) ou d'autres membres de la tribu Triticeae (par exemple Sr31 de seigle et Sr44 de Thinopyrum intermedium ).
Aucun des gènes Sr n'offre de résistance à toutes les races de rouille de la tige. Par exemple, beaucoup d'entre eux sont inefficaces contre la lignée Ug99 . Notamment Ug99 a la virulence contre Sr31, qui était efficace contre toutes les races de rouille de tige précédentes. Récemment, un nouveau gène de résistance à la rouille des tiges Sr59 de Secale cereale a été introgressées dans le blé, ce qui constitue un atout supplémentaire pour l'amélioration du blé afin d'atténuer les pertes de rendement causées par la rouille des tiges. Singh et coll. (2011) fournissent une liste des gènes Sr connus et leur efficacité contre Ug99.
Il existe une variation considérable entre les allèles Sr22, certains conférant une résistance et une certaine sensibilité.
Les agriculteurs éthiopiens ont largement adopté des variétés de blé résistantes depuis 2014.
Histoire de la rouille de la tige
Les ancêtres fongiques de la rouille noire infectent les graminées depuis des millions d'années et les cultures de blé aussi longtemps qu'elles sont cultivées. Selon Jim Peterson, professeur de sélection du blé et de génétique à l' Oregon State University , «la rouille des tiges a détruit plusieurs fois plus de 20% des récoltes de blé américaines entre 1917 et 1935 et les pertes ont atteint 9% deux fois dans les années 1950», avec la dernière épidémie aux États-Unis en 1962 détruisant 5,2% de la récolte.
Bien que l'Ug99 n'ait été découvert qu'en 1999, la rouille des tiges est un problème permanent qui remonte à l'époque d' Aristote (384-322 avant JC). Une pratique ancienne précoce par les Romains était celle où ils sacrifieraient des animaux rouges tels que des renards, des chiens et des vaches à Robigus ( fem. Robigo ), le dieu de la rouille. Ils effectueraient ce rituel au printemps lors d'un festival connu sous le nom de Robigalia dans l'espoir que la récolte de blé soit épargnée par la destruction causée par la rouille. Les enregistrements météorologiques de cette époque ont été réexaminés et il a été spéculé que la chute de l'Empire romain était due à une série de saisons des pluies dans lesquelles la rouille aurait été plus dure, entraînant des récoltes de blé réduites. Les lois interdisant l'épine-vinette ont été établies en 1660 à Rouen , France . Cela était dû au fait que les agriculteurs européens ont remarqué une corrélation entre l'épidémie d'épine-vinette et de rouille noire du blé. La loi interdisait la plantation d'épine-vinette près des champs de blé et était la première du genre.
Plus tard, la nature parasite de la rouille noire a été découverte dans les années 1700. Deux scientifiques italiens, Fontana et Tozzetti , ont d'abord expliqué le champignon de la rouille de la tige dans le blé en 1767. Le scientifique italien Giuseppe Maria Giovene (1753–1837), dans son travail Lettera al dottor Cosimo Moschettini sulla ruggine, a également étudié à fond la rouille de la tige. Trente ans plus tard, il a reçu son nom, Puccinia graminis , par Persoon et en 1854 les frères Louis René et Charles Tulasne ont découvert le stade caractéristique à cinq spores qui est connu dans certaines espèces de rouille de tige. Les frères ont également pu établir une connexion entre les spores rouges (urédospore) et noires (téliospore) en tant que différentes étapes du cycle de vie du même organisme, mais le reste des étapes est resté inconnu.
Anton de Bary a ensuite mené des expériences pour observer les croyances des agriculteurs européens concernant la relation entre les plants de rouille et d'épine-vinette, et après des tentatives réussies de relier les basidiospores du stade basidia à l'épine-vinette, il a également identifié que les éciospores au stade écie réinfectaient le hôte de blé. Après la découverte par de Bary des cinq stades de spores et de leur besoin d'épine-vinette en tant qu'hôte, John Craigie , un pathologiste canadien, a identifié la fonction du spermogonium en 1927.
En raison de la nature utile des plants d'épine-vinette et de blé, ils ont finalement été amenés en Amérique du Nord par des colons européens. L' épine-vinette a été utilisée pour beaucoup de choses comme faire du vin et des confitures des baies aux manches d'outils du bois. Finalement, comme ils l'ont fait en Europe, les colons ont commencé à remarquer une relation entre l'épine-vinette et les épidémies de rouille de la tige dans le blé. Des lois ont été promulguées dans de nombreuses colonies de la Nouvelle-Angleterre , mais lorsque les agriculteurs se sont déplacés vers l'ouest, le problème de la rouille des tiges s'est déplacé avec eux et a commencé à se propager dans de nombreuses régions, créant une épidémie dévastatrice en 1916. Ce n'est que lorsque deux ans plus tard, en 1918, que les États-Unis créé un programme pour supprimer l'épine-vinette. Le programme a été soutenu par des entités étatiques et fédérales et a été en partie motivé par la menace qu'il représentait pour les approvisionnements alimentaires pendant la guerre . La «guerre contre les barbaries» a été menée et a fait appel à l'aide des citoyens par des publicités à la radio et dans les journaux, des brochures et des kiosques justes demandant de l'aide à tous dans la tentative de débarrasser les arbustes d'épine-vinette de leur existence. Plus tard, en 1975–1980, le programme a été rétabli sous la juridiction d'état. Une fois que cela s'est produit, une quarantaine fédérale a été établie contre la vente d'épine-vinette sensible à la rouille des tiges dans les États qui faisaient partie du programme. Un programme d'analyse de l'épine-vinette a été créé pour s'assurer que seules les espèces et les variétés d'épine-vinette qui sont immunisées contre la rouille noire seront cultivées dans la zone de quarantaine.
En 1969, deux races non détectées auparavant en Australie ont été trouvées et pendant des décennies, une hypothèse était d' origine africaine, et en 2018, l'analyse ADN l'a confirmé spécifiquement sud-africain .
L'Afrique du Sud elle-même a un problème permanent avec diverses épidémies de rouille des tiges qui nécessitent une meilleure réponse, y compris un programme de sélection indigène pour la résistance .
Date de dernière mise à jour : 19/04/2023
Ajouter un commentaire