
Geosiphon pyriforme.
Geosiphon pyriforme
Geosiphon F.Wettst. (1915) est un genre de champignons de la famille des Géosiphonacées avec une seule espèce connue, Geosiphon pyriforme (Kotz.) F. Étoile. (1915). Le Geosiphon est le seul représentant de la classe des Gloméromycètes qui ne forme pas de mycorhizes arbusculaires avec des plantes supérieures, mais il forme un type de symbiose endosymique avec la cyanobacteria Nostoc ponctiform Hariot (1891).
Cet organisme a un micellio souterrain de croissance horizontale formé par des hyphes hautement ramifiés, des anaptosats et des cénocytaires qui forment de grandes clamidospores. Lorsque ces hyphes sont en contact avec une célula cellule de Nostoc punctiforme, avec laquelle elle partage un habitat, un processus d'endocytose commence qui culmine avec la formation d'une vésicule biliaire pyriforme multinucléé de 2 mm de long qui contient des filaments de cyanobactéries. À l'intérieur de cette vésicule biliaire, Nostoc effectue la photosynthèse à des vitesses plus élevées que celles qu'il présente dans la vie libre bénéficiant du champignon de la production ultérieure de sucres. Dans le même temps, les cyanobactéries prolifèrent nettement grâce aux phosphates que le champignon lui fournit et qui sont à très faibles concentrations dans l'environnement extérieur.[1]
La nature de cette association est problématique. D'une part, cet organisme est considéré par de nombreux auteurs comme un lichen comme une simple symbiose entre un champignon et une cyanobactérie. Compte tenu de la diversité des lichens connus et de leur origine paraphylétique, il est difficile de marquer plus de caractères communs à tous ces organismes. D'autre part, certaines définitions plus complexes de ce qu'un lichen est ne reconnaissent pas ces organismes comme une simple association, mais doivent avoir lieu avec les deux symbiotes en contact étroit. Il serait donc nécessaire d'avoir la présence d'une organisation des symbiotes dans des strates liées intimes, de structures de reproduction avec la présence des deux organismes tels que les isidiums ou les soredios ou des structures trophiques telles que les haussages du champignon pénétrant les cellules du photobiote. Étant donné que Geosiphon ne répond pas à ce critère de complexité, certains auteurs préfèrent appeler ce consortium endocitobiotique.
|
Geosiphon |
|
|
|
|
Royaume : |
|
|
Division : |
|
|
Service : |
|
|
Ordre : |
|
|
Famille : |
|
|
Geosiphon |
|
|
Geosiphon pyriforme |
|
{kind=link}
Description
{kind=link}
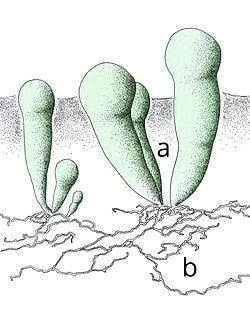
Anatomie de G. pyriforme.
{kind=link}
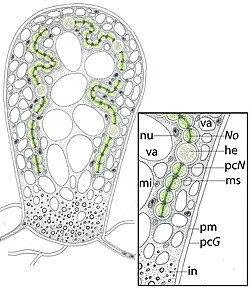
Structure interne de la vésicule biliaire de Geosiphon pyriforme.
Légende pc N : paroi cellulaire de Nostoc, No. : filament de Nostococ, ha : heterocisto, ms. : membrane simbiosomale, pm : plasmalem, pc G : paroi cellulaire de Geosiphon, n: nu : nu, mi. Mitochondria, dans.: inclusions citopasmatiques, va .
Cet organisme est un consortium endosimbiobiotique formé d'un champignon gingomique ou micosimbionte, Geosiphon pyriforme proprement dit, et d'une cyanobactérie filamenteuse ou photobiotique du genre Nostoc.
Dans le Geosiphon pyriforme, le champignon forme un myel a hypogueu et plagiotrophe (de croissance horizontale) formé par une ramification abondante, des anaptomes, des cénocytaires et non septées ou faiblement séparénés en maturité et en sénescence. Des groupes uniques ou petits sont traités dans la zone terminale de l'hypha, des mono-souldopors. Ces spores sont grandes, entre 160 et 320 m de diamètre et de blanc opaque, et rappellent morphologiquement celles produites par d'autres champignons mycorhizes arbusculaires avec des plantes supérieures.
En divers points du mycélium, les hyphes se développent verticalement à la surface du substrat formant un pyricle vert intense vestique, de long de 1 à 2 mm et d'environ 0,3 mm de diamètre, qui contient à l'intérieur des filaments de cyanobactéries qui synthétiseront les sucres via la photosynthèse pour le consortium.
Le filament de Nostoc est à son tour formé de cellules ovariennes sphériques qui ont généralement une constriction dans leur zone médiane et un diamètre de 0,5 m.[ ]Le champignon est capable de former des vésicules symbiotiques pyriformes toutes les trois semaines et chacun d'eux est fonctionnel pendant environ six mois, après quoi ils dégénèrent.
La vésicule biliaire est composée d'une membrane de chitine frébulaire épaisse, élastique et transparente qui maintient une pression interne proche des 6 bars. La matrice de la vésicule est formée principalement par un matériau muéculeux correspondent au cyplasme du champignon, des organites de la souris (dont un grand nombre de noyaux et de vacuoles) et par le simbiosme, une seule compartimentation formée par une membrane d'origine fongique contenant les filaments et les hétérocystes de Nostoc.
L'espace délimité par la membrane du symbiosome et de la paroi cellulaire des cyanobactéries, dit espace smbiosomal, a une épaisseur comprise entre 30 et 40 nm où il y a des cellules de Nostoc et entre 20 et 30 nm dans les endroits où elles n'ont pas. L'espace symbiosomal est occupé par un matériau amorphe qui semble être formé uniquement par le champignon puisqu'il n'a pas détecté la présence de mains, fucosa, N-acétylactosamine, acide sialique ou N-Acétilglucosamine, composants qui font partie de la couche de mucilage que Nostoc possède dans la vie libre.[6]
Le symbiosome contenant les filaments occupe principalement la zone périphérique des deux tiers supérieurs de la vésicule biliaire ainsi que de petites vacuoles fongiques et des mitochondries tandis que la zone interne se compose presque exclusivement de grands mucilages et de vaculums. Les cyanobactéries incluses dans ce consortium développent une taille cellulaire plus grande et une plus grande concentration de pigments que celle présentée par les mêmes organismes dans la vie libre ou dans d'autres associations symbiotiques. Le tiers inférieur de la vésicule est occupé par un mélange complexe de matrice, des granules de glycogène probablement formés par le Nostoc, [7]des vacuoles lipidiques, des microsomes et d'autres inclusions résultant metabolismodu métabolisme du champignon.
Formation de symbiose Geosiphon-Nostoc
Reconnaissance du photosym.

{kind=link}
Les fichiers de Nostoc avec hétérocèses.
La symbiose commence lorsque la fin de l'un des hyphes du champignon prend contact avec Nostoc, avec lequel il partage l'habitat. Les cyanobactéries qui forment la symbiose avec le Geosiphon sont le phanète Nostoc, un organisme capable de former une symbiose avec des plantes des genres Anthoceros, Blasia et Gunnera. Nostoc, contrairement à la plupart des algues participant à l'endosimbiose comme Cyanophora paradoxa, Glaucosphaera vacuolata, Gloeochaete wittrockiana ou Glaucocystis nostostochinearum, peuvent se développer indépendamment d'un macrobionte. Le naptochet est donc un symbiote optionnel tandis que le géosiphonpyriforme est un symbiote strict puisqu'il n'a pas été observé à ce jour de croissance sans former de symbiose.[9]Il existe une certaine spécificité dans l'association avec le mycobiotine au niveau moléculaire car seules certaines souches de Nostoc punctiform peuvent être incorporées par le champignon et, parmi celles-ci, quelques-unes permettent le développement du vésique symbiotique. Selon certaines recherches, les souches de Nostoc capables de former une symbiose sont celles qui faisaient auparavant partie d'une symbiose avec une plante de type Anthoceros ou Blasia.
Endosimbiose
Pour que le champignon ait lieu, les cyanobactéries doivent être dans un état de primordium, une forme dormante, car les hormogons mobiles ne sont jamais incorporés. Il est possible que l'élément clé de la reconnaissance des cyanobactéries par le champignon soit la présence dans la membrane des cellules du primordium du manoalaine polysaccharidique, absentes dans les hétérocistes et les acinètes. Le mécanisme, cependant, semble plus complexe car il a été observé que le champignon immobilise d'abord un hormogonio de Nostoc favorisant la transformation de celui-ci en primordium.
Une fois que Nostoc est immobilisé et dans le primordium, les cellules terminales du champignon hyphas excréent le cytoplasme sur les cellules des cyanobactéries en tant qu'étape avant leur endocytose. Cette endocytose dure entre 1 et 2 jours et semble avoir lieu par la restauration de la matière nucléagineuse excrétée, avec les cyanobactéries incorporées dans celle-ci, par les cellules terminales d'un ou plusieurs hyphes de. Après cela, les hyphes contenus par Nostoc et certains adjacents fusionnent leurs parois cellulaires et modifient leur morphologie en prenant une forme pyriforme pour former une vésicule multinucléique plus grande. La croissance ultérieure des hif fongiques maintient Nostoc dans cette structure toujours laissée à l'écart des hétéronistes.
Mature du symbiote.
Le stress causé dans les cellules cyanobactériennes qui ont été erthrompés leur fait perdre une partie de leurs pigments aux premiers stades de la symbiose. Cependant, immédiatement dans ce processus, lorsque le champignon commence à former la vésicule biliaire, les cyanobactéries récupèrent leur activité en synthétisant plus de pigments et en proliférant par division à un rythme compris entre 2 et 3 fois plus élevé que celui de l'organisme vivant en liberté.
Les cyanobactéries intégrées deviennent métaboliquement actives dans les vésicules qui acquièrent une couleur vert foncé et peuvent être observées à la surface du sol, exposées à la lumière du soleil. Les vésicules présentes dans leurs membranes de chitine ainsi que la membrane cellulaire des hyphages fongiques bien que leurs propriétés de perméabilité soient différentes de celles-ci et plus liées à celles des structures de la brousse de champignons mycorhizelics.
Avantages de la symbiose.
Une fois la symbiose formée, les cyanobactéries sont capables de fixer le CO 2 atmosphérique à l'intérieur des vésicules biliaires tandis que la fixation de l'azote se produit sur la partie des hétéronistes qui se sont formées dans le filament. L'incorporation de CO 2 est effectuée à des vitesses plus élevées en présence de la lumière au moyen de la voie fluorée phosphate qui dans l'obscurité au moyen de la voie carboxylase phosphoolpiruté comme c'est le cas dans les filaments de vie libre. Dans tous les cas, le taux total de fixation du CO 2 de Nostoc en symbiose est supérieur à celui qu'il possède en libre vie alors que la fixation de l'azote est plus faible dans les hétérofilis de dans l'endosimbiose avec le Geosiphon que dans ceux de Nostoc mycoshzial avec des plantes supérieures de sorte qu'il est intuitif la fonction principale des cyanobactéries dans cette symbiose est la photosynthèse.
La symbiose formée est bénéfique pour le champignon en obtenant une source endogène de sucres synthétisées par des cyanobactéries. Cet avantage est encore plus grand dans ce champignon car sa membrane externe est particulièrement imperméable à l'eau pour les molécules supérieures à 0,45 ou 0,5 nm, ce qui exclut par exemple l'incorporation de glucose du milieu extérieur.]Les, pour sa part, reçoivent des conditions d'homéostasie qui favorisent sa croissance et un complément de phosphate qui ne se trouve pas dans l'environnement extérieur.
Répartition et habitat

{kind=link}
Anthoceros agresstis.

{kind=link}
Blasia intervient.
Le Geosiphon pyriforme se développe dans la couche moins profonde de sols humides, argileux et polluants, restant son mycélium caché dans les premiers millimètres de substrat.Partager l'habitat avec une communauté de brophiles caractérisée par la présence de foies du genre Blasia et d'anthocérophytes du genre Anthoceros, organismes qui maintiennent également des relations symbiose avec Nostoc.
Bien que sa distribution soit censée être globale depuis la découverte de cette espèce, sa présence n'a été citée que dans cinq localités, toutes présentant les mêmes caractéristiques écologiques. Cependant, il ne peut actuellement être trouvé que dans l'un d'entre eux, situé dans les montagnes de Spessart, près de Bibergem-nd (Allemagne).
Filogenèse et taxonomies
Le nom générique, Geosiphon, vient des mots grecs ge () signifiant sol et sipho, siphonis () qui signifie tube.
L'épithète spécifique, pyriforme, vient des mots latinas latins pyrus, pyrus qui signifie poire et formule qui signifie forme. Plusieurs auteurs ont noté que la forme correcte de cette épithète devrait être pyriforme puisque le nom qui précède qu'il est mâle parce que c'est le mot sipho, siphonis.
Étant donné la complexité de cet organisme, son nom scientifique a d'abord été utilisé pour ne nommer que le symbiote, à partir de la conception qu'il s'agissait d'un seul organisme appartenant au groupe de lichens, puis jusqu'à aujourd'hui seulement pour le champignon, en maintenant le nom de « symbiose Geosiphon » ou .y-symbiosis Geosiphon-Nostoc.
Le pyriforme Geosiphon a été découvert en 1862 par Friedrich Traugott K.tzing et décrit à l'origine comme une algues siphonales nommée Botrydium pyiriforme. La découverte de la nature symbiotique de l'organisme n'a eu lieu qu'en 1915 Fritz von Wettstein a identifié la présence dans l'exploitation forestière d'un hôte ou d'un macrobiote, d'une algues siphonales hétérotrophes et d'une cyanobactérie bien connue, Nostoc. La véritable nature de l'hôte principal a finalement été reconnue en 1933 par Edgar Knapp en identifiant le macrosimbionte comme un champignon et en essayant de cultiver et de reproduire l'abattage de la symbion à partir de ses composants infructueux. Ce processus de culture a finalement été réalisé par Dieter Mollenhauer et Rosi Mollenhauer qui, en 1988, ont réussi à obtenir la taille complète du consortium par un processus complexe dans lequel des spores du champignon obtenues dans leur environnement naturel ont été contactées et qui ne pouvaient en soi former un mycélium avec des cultures de Nostoc filamenteux dans des moyens à très faible teneur en phosphate pour éviter la prolifération excessive des algues.
Les études filogenéticosphylogénétiques basées sur les petites sous-unités de l'ARN ont permis d'établir une relation phylogénétique entre le champignon Geosiphon et d'autres champignons mycorhizes arbusculaires tous dans la classe des Gloméromycètes.Malgré cette relation, il n'a pas été possible d'observer la présence de Geosiphon dans la micorrhiza de type arbusculaire avec une plante supérieure ou la formation d'endosymbiose avec des cyanocactères ou d'autres organismes photosynthétiques par tout gloméromite à l'exception de.
Date de dernière mise à jour : 29/06/2025
Ajouter un commentaire