
ECTOMYCORRHYZAE.
ECTOMYCORRHYZAE
Une ectomycorhize (de grec - ektos, "outside", mykes"gmonus", et "fungus", et rhiza"raciza", "racine"; pl. ectomycorrhizas ou ectomycorrhizae, abrégé EcM) est une forme de relation symbiotique qui se produit entre un symont fongique, ou mycobiont, et les racines de diverses espèces végétales. Le mycobiont provient souvent de la phyla Basidiomycota et d'Ascomycota, et plus rarement de la zygomycota. Les icolicorhizes se forment sur les racines d'environ 2 % des espèces végétales, généralement des plantes ligneuses, y compris des espèces de bouleaux, de diptérocarpe, de myrte, de hêtre, beechde saule, de pins et de roses. La recherche sur les ectomycorhizes est de plus en plus importante dans des domaines tels que la gestion et la restauration des écosystèmes, la sylviculture et l'agriculture.
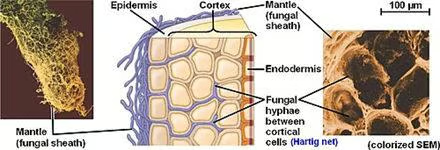
Contrairement à d'autres relations mycorhisiennes, telles que la mycorhize arbusculaire et la mycorhize ricoide, les champignons ectomycorhiziens ne pénètrent pas dans les parois cellulaires de leur hôte. Au lieu de cela, ils forment une interface entièrement intercellulaire connue sous le nom de filet de Hartig, composée d'hyphes fortement ramifiés formant un réseau entre les cellules épidermiques et les cellules racinaires corticales.
Les cécités sont en outre différenciées des autres mycorhizes par la formation d'une gaine dense d'hyphe, connue sous le nom de manteau, entourant la surface des racines. Ce manteau de gaine peut avoir jusqu'à 40 micromètres d'épaisseur, avec des hyphes s'étendant jusqu'à plusieurs centimètres dans le sol environnant. Le réseau d'hyphes aide la plante à absorber les nutriments, y compris l'eau et les minéraux, aidant souvent la plante hôte à survivre à des conditions défavorables .En échange, le symbionte fongique est doté d'un accès aux glucides.
Bien que des échantillons d'ectomycorhizes soient généralement prélevés à partir de l'horizon de surface en raison d'une densité de racines plus élevée, les ectomycorhizes sont connus pour se produire dans les racines des arbres profonds (une profondeur supérieure à 2 mètres), certains se produisant au moins jusqu'à 4 m.
Les corps de fructification fongiques EcM comprennent la truffe économiquement importante et comestible (Tuber) et les casquettes mortelles et les anges qui détruisent (Amanita).
Évolution
Les symbioses mycorhiziennes sont omniprésentes dans les écosystèmes terrestres, et il est possible que ces associations aient contribué à faciliter la colonisation des terres par les plantes. Il existe des preuves paléobinologiques et moléculaires que les mycorhizes arbusculaires (MA) sont apparues il y a au moins 460 millions d'années.
Les plantes et les champignons EcM présentent une large distribution taxonomique à travers tous les continents (en dehors de l'Antarctique), suggérant que la symbiose de l'EcM a d'anciennes racines évolutionnaires. Les pinacées sont la plus ancienne famille de plantes existantes dans laquelle se produit la symbiose avec les champignons EcM, et les fossiles de cette famille remontent à 156 millions d'années.
Il a été proposé que le type d'habitat et les fonctions distinctes des différentes mycorhémies aident à déterminer quel type de symbiose est prédominant dans une zone donnée. Dans cette théorie, les symbioses de l'EcM ont évolué dans des écosystèmes tels que les forêts boréales qui sont relativement productives mais dans lesquels le cycle des nutriments est encore en cours. Les cétomycorhizes sont intermédiaires dans leur capacité à absorber les nutriments, étant plus efficaces que les mycorhizes arbusculaires et moins que les mycorhizes ricoides, ce qui les rend utiles dans une situation de nutriments intermédiaire.
Paléobiologie
Les champignons sont composés de tissus mous, ce qui rend la fossilisation difficile et la découverte de fossiles fongiques rare. Cependant, certains spécimens exquisement conservés ont été découverts dans l'Éoccène Princeton Chert de la Colombie-Britannique. Ces fossiles ectomycorhiziens montrent clairement la preuve d'un filet, d'un manteau et d'hyphes de Hartig, démontrant des associations EcM bien établies il y a au moins 50 millions d'années.
Les fossiles montrent que les mycorhizes arbusculaires les plus courantes se sont formées bien avant d'autres types de symbioses de plantes fongiques. Les icactomycorhizes peuvent avoir évolué avec la diversification des plantes et l'évolution des conifères et des angiospermes. Les mycorhizes arbusculaires peuvent donc avoir été une force motrice dans la colonisation végétale de la terre, tandis que les ectomycorhizes peuvent avoir vu le jour soit en réponse à une spéciation plus poussée à mesure que le climat de la Terre est devenu plus saisonnier et plus aride, ou peut-être simplement en réponse à des habitats déficients sur le plan nutritionnel.
Études moléculaires
Les analyses moléculaires et phylogénétiques des linéies fongiques suggèrent que les champignons EcM ont évolué et ont persisté à de nombreuses reprises à partir d'ascendance non-EcM telles que l'humus et les champignons saprotrophes du bois. Les estimations vont de 7 à 16. Certaines études suggèrent qu'il y a eu des retours en situation de vie ancestrale, mais c'est controversé.
Morphologie
{kind=link}
Gradologie de base d'une association ectomycorhizienne commune
Comme suggéré par le nom, la biomasse des mycosmbionts est principalement extérieure à la racine de la plante. La structure fongique est composée principalement de trois parties: 1) les hyphes intraradicalaires constituent le filet de Hartig; 2) le manteau qui forme une gaine entourant la pointe de la racine; et 3) les hyphes extraradicalaires et les structures connexes qui se sont répandues dans tout le sol.
Net de Hartig
Le filet d'Hartig est formé par une croissance d'hyphes (souvent provenant de la partie interne du manteau environnant) dans la racine de l'hôte de la plante. Les hyphes pénètrent et croissent dans une direction transversale à l'axe de la racine, et forment ainsi un réseau entre les cellules externes de l'axe radiculaire. Dans cette région, les cellules fongiques et les cellules racinaires se touchent, et c'est là que se produit l'échange de nutriments et de carbone.
La profondeur de pénétration varie d'une espèce à l'autre. Dans l'eucalyptus et l'Alnus, le filet de Hartig est confiné à l'épiderme, alors que dans la plupart des gymnospermes, les hyphes pénètrent plus profondément, dans les cellules corticales ou l'endoderme. Dans de nombreux types d'épidermes, l'élongation des cellules le long de l'épiderme se produit, augmentant le contact de surface entre les champignons et les cellules racinaires. La plupart des filets de type corticaux Hartig ne montrent pas cet allongement, suggérant différentes stratégies pour augmenter le contact de surface parmi les espèces.
Manifestement
Une gaine d'hyphe connue sous le nom de manteau, qui a souvent plus de biomasse que l'interface du filet d'Hartig, enveloppe la racine. La structure du manteau est variable, allant d'un réseau d'hyphes lâches à une disposition structurée et stratifiée de tissu. Souvent, ces couches ressemblent au tissu du parenchyme végétal et sont appelées pseudo-parenchymateuses.
Parce que la racine est enveloppée par le manteau, elle est souvent affectée au développement. Les partenaires fongiques de l'EcM suppriment de manière caractéristique le développement des coiffes de leurs plantes. Ils peuvent également augmenter la ramification des racines en induisant des cytokines dans la plante. Ces motifs de ramification peuvent devenir si étendus qu'un seul manteau consolidé peut envelopper de nombreuses extrémités à la fois. Des structures comme celle-ci sont appelées ectomycorhizes tuberculés ou coralloides.
Les manteaux de différentes paires EcM présentent souvent des traits caractéristiques tels que la couleur, l'étendue de la ramification et le degré de complexité qui sont utilisés pour aider à identifier le champignon, souvent en tandem avec des analyses moléculaires. Les corps fruitiers sont également utiles mais ne sont pas toujours disponibles.
Hyphas extraradical et liaison
Plus d'informations : Mycélium extramatrique fulocmycorique

{kind=link}
Mycélium extraradical (blanc) sur les racines de Picea glauca (marron)
Les hyphes extraradiques s'étendent vers l'extérieur du manteau dans le sol, compensant la suppression des poils racinaires en augmentant la surface effective de la racine colonisée. Ces hyphes peuvent s'étaler seuls, ou dans un arrangement global connu sous le nom de rhizomorphe. Ces organes hyphal composites peuvent avoir une large gamme de structures. Certains rhizomorphes sont simplement des collections linéaires parallèles d'hyphes. D'autres ont une organisation plus complexe, par exemple les hyphes centraux peuvent être plus grands en diamètre que d'autres hyphes, ou les hyphes peuvent croître continuellement à la pointe, pénétrant dans de nouvelles zones d'une manière qui ressemble superficiellement à l'activité méristémique.
Cette partie de l'ectomycorhize, appelée mycélium extraradical ou extramatrique, fonctionne en grande partie comme une structure de transport. Ils s'étendent souvent sur des distances considérables, maintenant une grande zone de contact avec le sol. Certaines études ont montré une relation entre les taux de transport des nutriments et le degré d'organisation du rhizomorphe. Les rhizomorphes de différents types d'EcM ont souvent des types d'organisation et des stratégies d'exploration différents, observés comme une structure et une croissance différente au sein du sol. Ces différences aident également à identifier le champignon symbiotique.
Les hyphes s'étendant vers l'extérieur dans le sol à partir d'une ectomycorhize peuvent infecter d'autres plantes voisines. Les expériences et les études de terrain montrent que cela peut conduire à la formation de réseaux mycorhiziens communs (RMN) qui permettent le partage de carbone et de nutriments entre les plantes hôtes connectées. Par exemple, l'isotope rare 14 a été ajouté à un arbre particulier et plus détecté par la suite dans des plantes et des plants voisins. Une étude a observé un transfert de carbone bidirectionnel entre Betula papyrifera et Pseudotsuga menziesii, principalement à travers les hyphes de l'ecfomycorhize. Cependant, toutes les plantes ne sont pas compatibles avec tous les réseaux fongiques, de sorte que toutes les plantes ne peuvent pas exploiter les avantages des liens ectomycorhiziens établis.
Il a été suggéré que le raccordement commun des nutriments par les CMN s'implique dans d'autres processus écologiques tels que l'établissement de semis, la succession des forêts et d'autres interactions entre plantes et plantes. Il a été démontré que certaines mycorhizes arbusculaires transportent des signes d'alerte sur le réseau d'attaques par des insectes ou des maladies.
Corps fructifiants

{kind=link}
L'hypogée de la rotule de, la truffe noire du Périgord
Contrairement à la plupart des champignons mycorhiziens arbusculaires, les champignons EcM se reproduisent sexuellement et produisent des corps de fructification visibles sous une grande variété de formes. Le corps fructifiant, ou sporocarpe, peut être considéré comme une extension des hyphes extraradicalaires. Ses parois cellulaires et ses spores sont généralement composées de glucides complexes, et incorporent souvent une grande partie de l'azote. De nombreux champignons de l'EcM ne peuvent former des corps fructifiants et pas plus de cycle de vie en participant à une relation EcM.
Les corps fruitiers de nombreuses espèces prennent des formes classiques et bien reconnues telles que des epigeous champignons épigés et des truffes hypogéens. La plupart d'entre eux produisent des propagules microscopiques d'environ 10 pm qui peuvent se disperser sur de grandes distances par l'intermédiaire de divers vecteurs, allant du vent aux animaux mycophages. Il a été suggéré que les animaux sont attirés par des corps fructifiants hypogésiques parce qu'ils sont riches en nutriments tels que l'azote, le phosphore, les minéraux et les vitamines. Toutefois, d'autres font valoir que les nutriments spécifiques sont moins importants que la disponibilité des aliments à des moments précis de l'année.
Des enquêtes sur des organismes de fructification ont été utilisées pour évaluer la composition et la richesse des communautés dans de nombreuses études. Cependant, cette méthode est imparfaite car les corps fructifeurs ne durent pas longtemps et peuvent être difficiles à détecter.
Physiologie
Presymbiose
Pour former une connexion ectomycorhizienne, les hyphes fongiques doivent d'abord croître vers les racines de la plante. Ensuite, ils doivent s'envelopper et pénétrer dans les cellules de la coiffe et les infecter, permettant à la symbiotique Hartig net et aux structures associées de se former. Les deux partenaires (plantes et champignons) doivent suivre une séquence précise d'expression génique pour que cela soit réussi. Il est prouvé que la communication entre les partenaires au stade précoce de l'ectomycorhizige se produit dans certains cas au moyen de composés organiques volatils produits uniquement pendant la phase d'interaction, et que les gènes impliqués dans les processus de croissance sécrétrice, apicale et d'infection montrent des changements d'expression au début de la phase de pré-contact. Ainsi, un ensemble complexe de changements moléculaires semble avoir lieu avant même que le champignon et la plante hôte ne prennent contact.
Les hôtes des plantes libèrent des métabolites dans la rhizosphère qui peuvent déclencher la germination des basidiospores, la croissance des hyphes vers la racine et les premières étapes de la formation de l'EcM. Il s'agit notamment des flavonoides, des diterpènes, des cytokinines, des hormones et d'autres nutriments. Il a été démontré que certains métabolites libérés par l'hôte stimulent la croissance fongique de Pisolithus, modifient l'angle de ramification des hyphes et provoquent d'autres modifications du champignon. Certains gènes fongiques semblent être exprimés avant le contact des plantes, suggérant que les signaux dans le sol peuvent induire des gènes fongiques importants à distance de la plante.
Symbiose
Une fois que les hyphes fongiques entrent en contact avec les cellules de la coiffe des racines, ils doivent continuer à croître vers l'intérieur des cellules épidermiques et se multiplier pour former les couches qui finiront par produire le manteau. La production du manteau fongique implique la régulation à la hausse des gènes responsables de la traduction et de la croissance cellulaire, ainsi que des responsables de la synthèse et du fonctionnement membranaires, tels que les hydrophobes. Certains polypeptides ne sont trouvés que lorsque le champignon et la plante ont atteint la symbiose; ces protéines liées à la symbiose (SR) sont appelées ectomycorhitanines.
Les modifications majeures de la synthèse des polypeptides et des ARNm se produisent rapidement après la colonisation par le champignon, y compris la production d'ectomycorhizines. Les changements incluent la régulation à la hausse des gènes qui peuvent aider les nouvelles membranes à se former à l'interface symbiotique. L'effet du manteau sur la prolifération des racines, le développement des cheveux radiculaires et la ramification dichotomique peut être partiellement imité par les exsudats fongiques, fournissant un chemin pour identifier les molécules responsables de la communication.
Le filet de Hartig se forme initialement à partir de la couche intérieure complètement différenciée du manteau, et la pénétration se produit dans un large front orienté perpendiculairement à l'axe de la racine, en creusant à travers l'espace apoplasique. Certaines cellules végétales réagissent en produisant des protéines liées au stress et à la défense, y compris les chitinases et les peroxydases qui pourraient inhiber la formation du filet d'Hartig. Cependant, une colonisation étendue des racines se produit toujours dans ces plantes et ces caractéristiques de résistance semblent diminuer d'environ le jour 21 après la colonisation, ce qui implique que les champignons EcM peuvent supprimer la réponse de la défense.
Comme le champignon et la plante deviennent étroitement liés, ils commencent à partager les nutriments. Ce processus est également contrôlé par des gènes liés à la symbiose. Par exemple, la captation monosaccharide dans Amanita muscaria nécessite un transporteur qui n'est exprimé que lorsqu'il est dans une association mycorhizienne. Lorsque le transporteur est exprimé, ce qui entraîne une augmentation des importations de sucre par le champignon, l'hôte de la plante répond en augmentant la disponibilité du sucre. Le transport d'ammonium et d'acides aminés du champignon vers la plante est également réglementé.
Expansion et échange de nutriments
L'azote est essentiel dans la biochimie des plantes, étant nécessaire pour la chlorophylle et toutes les protéines. Dans la plupart des écosystèmes terrestres, l'azote est rare et est piégé dans la matière organique qu'il est difficile de décomposer. Les symbiotes fongiques offrent ainsi deux avantages aux plantes : la plus grande gamme de leurs hyphes par rapport aux racines, et une plus grande capacité à extraire l'azote de la couche de sol dans laquelle se trouve la matière organique. Le transfert net de nutriments vers les plantes nécessite que le nutriment franchisse trois interfaces : 1) l'interface sol-fungus, 2) l'interface champignon-apoplaste, et 3) l'interface apoplaste-aspiration. Il a été estimé que les champignons ectomycorhizants reçoivent environ 15 % du produit alimentaire de la plante hôte et, en retour, fournissent jusqu'à 86 % des besoins en azote d'un hôte.
Certaines études ont montré que si l'on dispose d'une quantité excessive d'azote disponible en raison de l'utilisation humaine d'engrais, les plantes peuvent détourner leurs ressources du réseau fongique. Cela peut poser des problèmes pour le champignon, qui peut être incapable de produire des corps fructifiants, et à long terme peut entraîner des changements dans les types d'espèces fongiques présentes dans le sol. Dans une étude, la richesse des espèces a diminué de façon spectaculaire avec l'augmentation des apports d'azote, plus de 30 espèces étant représentées sur des sites à faible teneur en azote et seulement 9 sur des sites à forte teneur en azote.
Comme les hyphes de la région du filet de Hartig deviennent de plus en plus emballés, ils s'appuient contre les parois cellulaires des cellules racinaires de la plante. Souvent, les parois fongiques et végétales deviennent presque indiscernables là où elles se rencontrent, ce qui facilite le partage des nutriments. Dans de nombreuses ectomycorhizes, les hyphes de filet de Hartig manquent de divisions internes, créant une structure de transmission multinucléaire qui facilite le transport interhyphal. Les hyphes ont une forte concentration d'organites responsables de la production d'énergie et de protéines (mitochondria et réticulum endoplasmique rugueux) à leurs extrémités. Il existe des signes indiquant que les transporteurs dans les membranes plasmiques fongiques et végétales sont actifs, suggérant un échange bidirectionnel de nutriments.
{kind=link}
Ectomycorhize avec Douglas-fir (Pseudotsuga menziesii) et Cortinarius spp.
La structure du réseau EcM dépend de la disponibilité des nutriments. Lorsque la disponibilité des nutriments est faible, l'investissement dans le réseau souterrain est élevé par rapport à la croissance en sus du sol. Le phosphore est un autre qui limite les nutriments dans de nombreux écosystèmes terrestres. Les preuves suggèrent que le phosphore est transféré en grande partie sous forme d'orthophosphate. Certaines ectomycorhizes formant des tapis contiennent des ribonucléases capables de dégrader rapidement l'ADN pour obtenir du phosphore à partir de noyaux.
Prestations non nutritionnelles
Les hyphes extraradiques, en particulier les rhizomorphes, peuvent également offrir un transport inestimable de l'eau. Souvent, ils se développent en coureurs spécialisés qui s'étendent loin des racines de l'hôte, augmentant la zone d'accès à l'eau fonctionnelle. La gaine d'hyphe enveloppant les extrémités des racines agit également comme une barrière physique protégeant les tissus végétaux des agents pathogènes et des prédateurs. Il existe également des preuves que les métabolites secondaires produits par les champignons agissent comme des mécanismes de défense biochimiques contre les champignons pathogènes, les nématodes et les bactéries qui pourraient tenter d'infecter la racine mycorhizienne. De nombreuses études montrent également que les champignons EcM permettent aux plantes de tolérance des sols à des concentrations élevées de métaux lourds, des sels et des polluants organiques.
Ectendomycorhizes
Bien que le filet Hartig se forme en dehors des cellules racinaires, la pénétration des cellules corticales des plantes se produit occasionnellement. De nombreuses espèces de champignons ectomycorhiziens peuvent fonctionner soit comme des ectomycorhizes, soit dans le mode pénétrant typique des mycorhizes arbusculaires, selon l'hôte. Ces associations représentant une forme de symbiose entre les mycorhizes arbusculaires et les ectomycorhizes, elles sont appelées ectendomycorhizes.
Écologie
Biogéographie et gradients environnementaux
Les champignons eredyhonsiens se trouvent dans tous les écosystèmes boréaux, tempérés et tropicaux, principalement parmi les familles dominantes productrices de plantes ligneuses. Nombre des familles de champignons communes dans les forêts tempérées (ex. Les russulaceae, Boletae, Thelephoraceae) sont également répandues dans l'hémisphère sud et les forêts tropicales de diptérocarpe: bien que les familles de plantes soient très différentes dans les forêts tempérées et tropicales, les champignons ectomycorhiziens sont assez similaires. Les types de champignons EcM sont affectés par les types de sol à la fois dans le champ et dans le laboratoire.
Pour la plupart des types de plantes et d'animaux, la diversité des espèces augmente vers l'équateur. C'est ce qu'on appelle le gradient latitudinal de la diversité (LGD). En revanche, il est prouvé que les champignons EcM peuvent être à la plus grande diversité dans la zone tempérée. Si tel est le cas, cela pourrait s'expliquer par une ou plusieurs des hypothèses suivantes: 1) les champignons EcM peuvent avoir évolué à des latitudes plus élevées avec les hôtes des Pinacées, et être moins capables de rivaliser dans les climats tropicaux; 2) les plantes utilisées par les EcM comme hôtes pourraient être plus diverses dans les conditions tempérées, et la structure du sol dans les régions tempérées peut permettre une plus grande diversité.
Possibilité d'accueil et réponses communautaires
La plupart des hôtes de l'EcM présentent de faibles niveaux de spécificité, et peuvent former des symbioses avec de nombreux champignons éloignés. Cela peut avoir des avantages évolutifs pour la plante de deux manières : 1) les plants de la plante sont plus susceptibles d'être capables de former des mycorhizes dans un large éventail d’habitats ; et 2) la plante peut utiliser différents champignons qui varient dans leur capacité à accéder aux nutriments.
Les champignons EcM présentent différents niveaux de spécificité pour leurs hôtes de plantes, et les coûts et les avantages de leur spécialisation ne sont pas bien compris. Par exemple, le groupe suilloide, un assemblage monophylétique contenant les genres Suillus, Rhizopogon, Gomphidius et d'autres, montre un degré extrême de spécificité, avec la quasi-totalité de ses membres formant des ectomycorhizes avec des membres des Pinaceae. Cependant, de nombreux autres groupes de champignons présentent une très large gamme.
Les plantes hôtes qui sont taxonomiquement liées ont plus de communautés fongiques EcM similaires que les taxons qui sont plus liés à la distance. De même, les études phylogénétiques moléculaires ont montré que les champignons dérivés d'un ancêtre commun sont plus susceptibles d'avoir des hôtes taxonomiquement liés. La maturité de l'environnement hôte, ou son statut de succession, peut également affecter la variété des communautés fongiques EcM présentes. D'autres facteurs indirects peuvent également jouer un rôle dans la communauté fongique de l'EcM, tels que la chute des feuilles et la qualité de la litière, qui affectent les niveaux de calcium et le pH du sol.
Rôles dans l'invasion

{kind=link}
Plantation de pins, probablement inoculée avec des spores fongiques pour permettre la formation d'ectomycorhizes bénéfiques
Les plantes qui ne sont pas originaires d'une région ont souvent besoin de symbiotes mycorhiziens pour prospérer. La grande majorité des mycorhizes arbusculaires ne sont pas spécifiques, de sorte que les plantes qui interagissent avec ces mycorhizes deviennent souvent invasives rapidement et facilement. Cependant, les symbioses ectomycorhissiques sont souvent relativement spécifiques. Dans la foresterie exotique, des champignons EcM compatibles sont souvent introduits dans le paysage étranger pour assurer le succès des plantations forestières. C'est le plus fréquent chez les eucalyptes et les pins, qui sont des arbres ectomycorhiziens obligatoires dans des conditions naturelles. Les pins étaient difficiles à établir dans l'hémisphère sud pour cette raison, et de nombreuses plantations d'eucalyptus nécessitaient une inoculation par les champignons EcM à partir de leur paysage natal. Dans les deux cas, une fois les réseaux EcM introduits, les arbres ont pu se naturaliser et ont ensuite commencé à rivaliser avec les plantes indigènes.
Cependant, de nombreuses espèces d'EIM co-vavirent sans l'aide de l'activité humaine. Les Pinacées de la famille envahissent souvent les habitats ainsi que les champignons EcM spécifiques des genres Suillus et Rhizopogon. Il existe également des champignons formant des ectomycorhizes avec des distributions cosmopolites qui peuvent permettre aux espèces végétales non indigènes de se propager en l'absence de leurs champignons EcM spécifiques de l'écosystème indigène.
Les plantes peuvent rivaliser en attaquant les réseaux fongiques des uns et des autres. Les plantes indigènes dominantes peuvent inhiber les champignons EcM sur les racines des plantes voisines, et certaines plantes invasives peuvent inhiber la croissance des champignons ectomycorhiziens indigènes, en particulier s'ils s'établissent et dominants. La moutarde invasive à l'ail, Alliaria petiolata, et son isothiocyanate allélochimique de benzyle, ont été démontrés comme étant la croissance de trois espèces de champignons EcM cultivées sur des plants de pin blanc. Les changements dans les communautés de l'EcM peuvent avoir des effets dramatiques sur l'absorption des nutriments et la composition communautaire des arbres indigènes, avec des ramifications écologiques de grande portée.
Compétition et autres symbiontes végétales
La concurrence entre les champignons EcM est un cas bien documenté d'interactions microbiennes du sol. Dans certaines expériences, le moment de la colonisation par des champignons EcM concurrents a déterminé quelle espèce était dominante. De nombreux facteurs biotiques et abiotiques peuvent médier la concurrence entre les champignons EcM, tels que la température, le pH du sol, l'humidité du sol, la spécificité de l'hôte et le nombre de concurrents, et ces facteurs interagissent les uns avec les autres d'une manière complexe. Il existe également des preuves de la compétition entre les champignons EcM et les champignons mycorhiziens arbusculaires. Cela est surtout noté chez les espèces qui peuvent accueillir à la fois des champignons EcM et AM sur leurs racines.
Certaines bactéries du sol, connues sous le nom de bactéries d'aide à Mycorrhiza (MHBs), ont montré qu'elles stimulent la formation d'EcM, la biomasse des racines et des pousses et la croissance fongique. Certains font valoir que les bactéries de ce type devraient être considérées comme un troisième composant des mycorhhizes. D'autres bactéries inhibent la formation ectomycorhizienne.
Interactions avec les animaux

{kind=link}
Le corps de fructification épigé l'éventuel de Cantharellus cibius, la chanterelle d'or.
De nombreux champignons ectomycorhiziens s'appuient sur les mammifères pour la dispersion de leurs spores, en particulier les champignons avec des corps fructifiants hypogésiques. De nombreuses espèces de petits mammifères sont des mycophages, mangeant un large éventail de champignons et surtout les corps fructifiants. Les spores sont dispersées soit parce que le corps fructif est déterré et cassé, soit après ingestion et excrétion ultérieure. Certaines études suggèrent même que le passage à travers l'intestin d'un animal favorise la germination des spores, bien que pour la plupart des espèces fongiques, cela ne soit pas nécessaire. En propageant les spores fongiques, ces animaux ont un effet indirect sur la structure de la communauté des plantes.
D'autres corps fructifiants sont mangés par des invertébrés tels que les mollusques et les larves de mouches, dont certains sont même tolérants à la z-amanitine toxique trouvée dans les casquettes de mort. Au-dessous du sol, les nématodes et les écailles consomment également des tissus fongiques.[15]Le champignon ectomycorhizien Laccaria bicolor a été trouvé pour attirer et tuer les braillants de printemps pour obtenir de l'azote, dont certains peuvent ensuite être transférés à la plante hôte. Dans une étude, le pin blanc de l'Est inoculé avec L. bicolor a pu déduire jusqu'à 25 % de son azote des briques de printemps.[85]
Les champignons comestibles sont importants dans les sociétés du monde entier. Les truffes, les porcins et les chanterelles sont connus pour leur importance culinaire et financière.
Production végétale
Agriculture
Les champignons eredyhhiziens ne sont pas prédominants dans les systèmes agricoles et horticoles. La plupart des plantes cultivées économiquement pertinentes qui forment des mycorhizes ont tendance à les former avec des champignons mycorhiziens. De nombreuses pratiques agricoles modernes telles que le travail du sol, les engrais lourds et les fongicides sont extrêmement préjudiciables aux mycorhizes et à l'écosystème environnant. Il est possible que l'agriculture affecte indirectement les espèces et les habitats ectomycorhiziens à proximité ; par exemple, l'augmentation de la fécondation diminue la production de sporocarpes.
Foresterie
Dans le secteur forestier commercial, la transplantation d'arbres cultivés à de nouveaux endroits nécessite souvent un partenaire ectomycorhizien qui accompagne. Cela est particulièrement vrai pour les arbres qui ont un degré élevé de spécificité pour leur mycobionte, ou les arbres qui sont plantés loin de leur habitat d'origine parmi de nouvelles espèces fongiques. Cela a été montré à plusieurs reprises dans les plantations impliquant des arbres ectomycorhiziens obligatoires, tels que les espèces et de Pinus. La plantation de masse de ces espèces nécessite souvent un inoculum de champignons de l'EcM pour les arbres pour prospérer.
Parfois, des espèces de plantations ectomycorhiziennes, telles que le pin et l'eucalyptus, sont plantées et promues pour leur capacité à agir comme puits sinkde carbone atmosphérique. Cependant, les champignons ectomycorhiziens de ces espèces ont également tendance à épuiser le carbone du sol, rendant cette utilisation controversée des plantations.
Restauration
Le rôle des ectomycorhizes dans le soutien de leurs plantes hôtes a conduit à suggérer que les champignons EcM pourraient être utilisés dans des projets de restauration visant à rétablir les espèces végétales indigènes dans les écosystèmes perturbées par une variété de problèmes. Étant donné que la disparition de champignons mycorhizaux à partir d'un habitat constitue un événement majeur de perturbation du sol, leur ré-addition est un élément important de l'établissement de la végétation et de la restauration des habitats.
Résilience dans des environnements difficiles
Métaux lourds
Les métaux lourds sont toxiques pour les organismes vivants. Les concentrations élevées de métaux lourds tels que le zinc, le cuivre, le cadmium, le plomb, le nickel et le chrome affectent les processus métaboliques de base et peuvent entraîner des dommages cellulaires et la mort. Certains champignons ectomycorhizants sont tolérants aux métaux lourds, de nombreuses espèces ayant la capacité de coloniser des sols contaminés.]Il existe également des cas de populations localement adaptées pour tolérer des environnements chimiques difficiles.
Les champignons présentent des mécanismes de détoxification pour réduire les concentrations de métaux lourds dans leurs cellules. Ces mécanismes comprennent la réduction de l'absorption des métaux lourds, la séquestration et le stockage des métaux lourds dans la cellule, et l'excrétion. L'absorption des métaux lourds peut être réduite par sorption et inactivation métabolique au niveau de la paroi cellulaire et de l'applaste. Les champignons cétomycorhiziens ont également la capacité de lier des quantités considérables de métaux lourds. Une fois à l'intérieur de la cellule, les métaux lourds peuvent être immobilisés dans des complexes organo-métalliques, rendus solubles, transformés en métalloloniques, impliqués dans la séquestration des métaux et/ou stockés dans les vacuoles sous des formes chimiquement inactives. Des systèmes de détoxification antioxydant peuvent également être en place, réduisant la production de radicaux libres et protégeant la cellule fongique. Les champignons peuvent exporter les métaux du cytoplasme vers l'applaste, un mécanisme qui se produit également dans les plantes. Les champignons ectomycorhiziens peuvent également concentrer les métaux lourds dans leurs corps fructifeurs. Des différences génétiques entre les populations vivant dans des habitats toxiques ou non toxiques ont été rarement rapportées, ce qui indique que la tolérance aux métaux est généralisée. Aucun tax fiscal endémique adapté aux métaux n'a été documenté jusqu'à présent. Il existe toutefois des preuves concernant les déplacements des communautés associés aux métaux lourds, avec une plus faible diversité associée aux sites contaminés. En revanche, les sols naturellement riches en métaux lourds, tels que les sols serpentins, ne semblent pas affecter la diversité des communautés fongiques ectomycorhiziennes.

{kind=link}
Suillus lutéus, un champignon ectomycorhizien avec des écotypes connus pour être associés à des concentrations de métaux lourds
Bien que la tolérance généralisée aux métaux semble être la norme pour les champignons ectomycorhiziens, il a été suggéré que quelques champignons tels que Pisolithus tinctorius, P. albus et des espèces du genre Suillus peuvent s'adapter à des niveaux élevés d'Al, 'n, Cd et Cu. Suillus luteus et S. bovinus sont de bons exemples, avec des écotypes connus adaptés à zn, Cd et Cu.
Pollution et phytorelogie
Il a été constaté que les champignons EcM ont des effets bénéfiques dans plusieurs types d'environnements pollués, notamment :
Sel riche : Un certain nombre d'études ont montré que certains champignons EcM peuvent aider leurs hôtes à survivre à des conditions de salinité élevées dans le sol.
- Radionucléides : De nombreuses espèces de champignons ectomycorhiziens, dont les Cortinariaceae, peuvent s'aggraver les radionucléides.
- Polluants organiques: Certaines espèces de la République de Chine méridionale sont capables de décomposer les polluants organiques persistants (POP) tels que les organochlorures et les polychlorobiphényles (PCB). Les produits chimiques qui peuvent être détoxifiés par les champignons EcM, seuls ou en association avec leur plante hôte, comprennent le 2,4-dichlorophénol et le tétrachloroéthylène.
Changements climatiques
Les communautés équatoriennes peuvent être touchées par l'augmentation du CO 2 et les effets du changement climatique qui en découlent. Dans certaines études, des taux élevés de CO 2 ont augmenté la croissance du mycélium fongique et une augmentation de la colonisation de la racine de l'EcM. D'autres associations de la MCE ont montré peu de réponse à une augmentation du CO 22.
L'augmentation des températures donne également une gamme de réponses, certaines négatives, et d'autres positives. La réponse de l'EcM à la sécheresse est complexe car de nombreuses espèces assurent une protection contre la dessiccation des racines et améliorent la capacité des racines à absorber l'eau. Ainsi, les EcM protègent leurs plantes hôtes en période de sécheresse, bien qu'elles puissent elles-mêmes être touchées au fil du temps.
Conservation
Alors que l'importance des organismes souterrains pour la productivité forestière, la récupération et la stabilité deviennent évidentes, la conservation des ectomycorhizes retient l'attention. De nombreuses espèces de champignons EcM en Europe ont diminué, en raison de facteurs tels que la diminution de la vitalité des arbres, la conversion des forêts en d'autres utilisations, la pollution et l'acidification des sols forestiers. Il a été soutenu que la conservation des ectomycorhizes nécessite une protection des espèces dans toute leur aire de répartition et leur habitat, [88]afin de s'assurer que tous les types de communautés EcM sont préservés.
Le Plan forestier du Nord-Ouest, qui régit l'utilisation des terres fédérales dans la région du nord-ouest du Pacifique aux États-Unis, comprend des dispositions pour l'étude des champignons menacés et l'élaboration de stratégies pour les gérer et les protéger. Le Conseil européen pour la conservation des champignons a été créé en 1985 pour promouvoir la recherche et l'attention sur les champignons en danger.[119]En 2018, le Conseil a collaboré avec les Jardins botaniques royaux de Kew pour produire le rapport sur l'état des champignons dans le monde, 2018.

{kind=link}
Brûlure prescrite dans un peuplement de Pinus nigra
Les stratégies de conservation comprennent l'entretien: 1) des plantes refuges et des hôtes réservoirs pour préserver la communauté fongique EcM après la récolte; 2) des arbres matures pour fournir aux plants un éventail varié de champignons EcM; et 3) des peuplements anciens qui ont divers macro- et micro-habitats et soutiennent des communautés fongiques EcM variées. La préservation des constituants naturels des sols forestiers et la rétention des débris et des substrats ligneux peuvent également être importantes. Dans une étude concernant les plants de sapins de Douglas, l'élimination des débris de sol forestier et le tassement du sol ont diminué la diversité et l'abondance des champignons EcM de 60 %[ ]L'enlèvement de l'herbe de pin a réduit de la même manière la diversité et la richesse des champignons EcM. Certaines stratégies, telles que les brûlures prescrites, ont des effets différents sur différents types de communautés EcM, allant de moins) à neutres ou positifs.
De grandes collections de champignons, y compris des champignons ectomycorhiziens, sont maintenues dans le monde entier en tant qu'assurance contre la perte génétique. Toutefois, ces collections sont incomplètes.
Date de dernière mise à jour : 22/07/2025
Ajouter un commentaire