
SYSTÉMATIQUE DES VÉGÉTAUX
1/ Les grands groupes végétaux
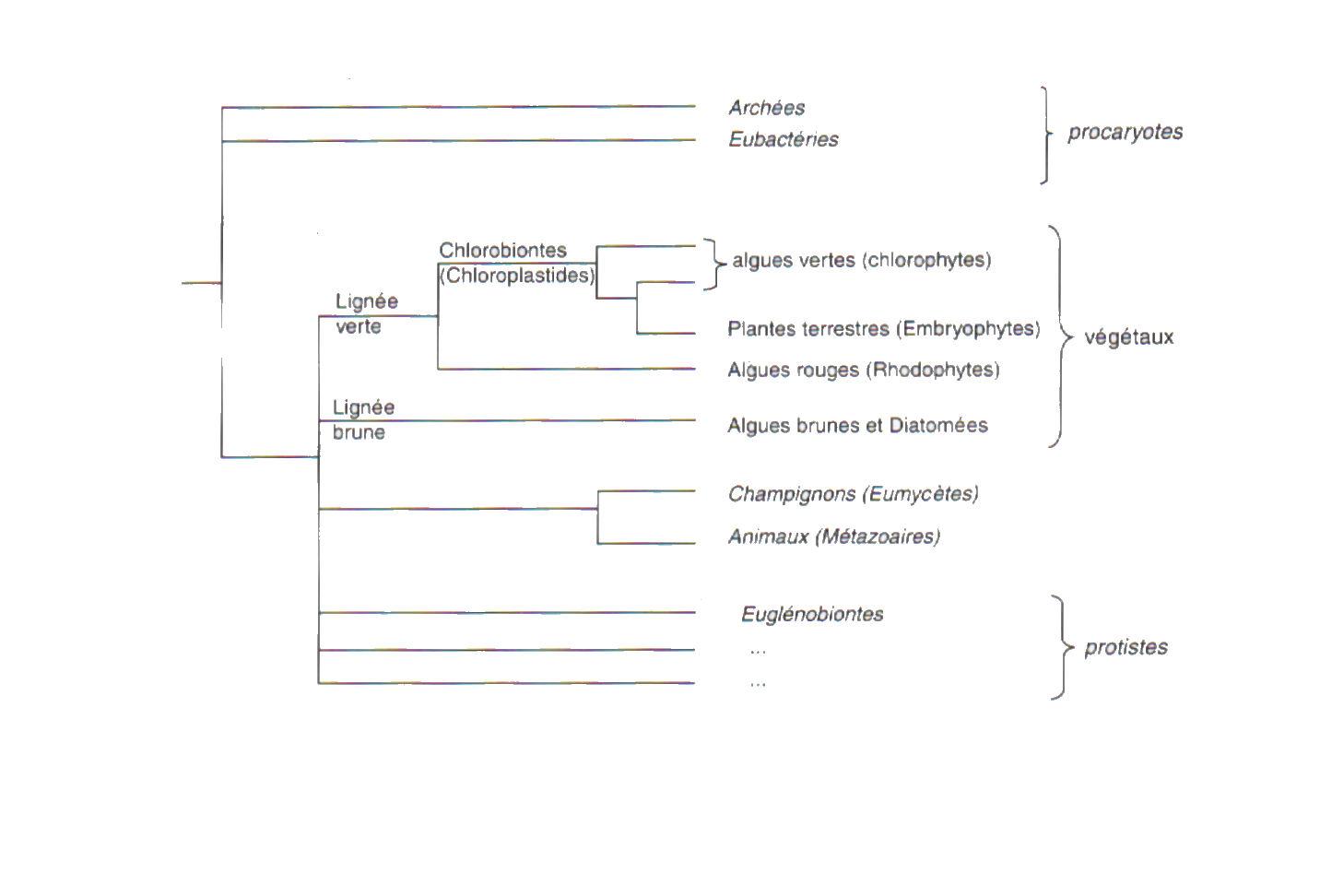
Dans la division du vivant en deux règnes – animaux et végétaux – bactéries et champignons faisaient partie des végétaux. Aujourd’hui six grands ensembles : Archées, Eubactérie, Protiste, Champignons (incluant les lichens), animaux et végétaux. Les deux premiers possèdent une cellule procaryote réduite à une paroi externe, une membrane plasmique, et un ADN circulaire inclus dans un cytoplasme. Les quatre ensembles suivants ont une cellule eucaryote pourvue d’une membrane plasmique protégée au nom par une paroi externe, d’un cytoplasme comportant un noyau, un cytosquette, un système endomembranaire, éventuellement des flagelle locomoteurs et dotée, par endosymbiose, d’organites intracellulaires.
L’incorporation de Protéobactéries (Eubactérie pourpres) par endosymbiose est à l’origine des mitochondries, organites présents chez l’ensemble des eucaryotes. L’incorporation, toujours par endosymbiose, de Cyanobactéries (Eubactéries photosynthétiques) est à l’origine des chloroplastes de la lignée verte. Cette lignée comprend les algues rouges (Rhodophytes ou Rhodobiontes) chez lesquelles la couleur verte de la chlorophylle est masquée par la phycoérythrine pigment rouge et les chlorobiontes, en plus de la chlorophylle a, par de la chlorophylle b.
Les chlorobiontes comprennent les algues vertes et les plantes terrestres.
Une nouvelle endosymbiose (appelé endosymbiose secondaire), par ingestion d’un eucaryote unicellulaire issu des Algues rouges par une autre cellule eucaryote, conduit à la lignée brune des Ochrophyta lesquels doivent leur couleur à la fucoxanthine, pigment accessoire brun. Certains des Ochrophyta ont perdu secondairement leurs pigments photosynthétiques et se comportent comme des « champignons ». De même l’endosymbiose secondaire d’un eucaryote unicellulaire issu des algues vertes par une autre cellule eucaryote conduit aux Euglénobiontes qui, elles aussi, peuvent perdre secondairement leur chlorophylle. La notion de végétal correspond à des Eucaryotes qui ont intégré directement ou indirectement, des procaryotes photosynthétiques même s’ils ont perdu secondairement leurs chloroplastes, comme chez les Oomycètes ou les angiospermes parasites.
Les champignons (ou Eumycètes) sont un groupe frère des choanoflagellés et des Éponges donc plus proches des animaux, que des végétaux. Quand aux lichens, ils résultent de la symbiose d’une Cyanobactérie ou d’une algue verte unicellulaire et d’un champignon.
2/ Embranchement, classe, ordre, famille, genre, espèce
Les grands groupes que nous venons de distinguer correspondent à des lignées évolutives différentes. Certains sont monophylétiques (du grec phylon, race, tribu) : chacun d’eux sont un clade (du grec klados, rameau). Par exemple, le clade des Chlorobiontes comprend toutes les plantes terrestres, ainsi que toutes les algues vertes et leurs ancêtres communs. Certaines entités, comme les algues, ne correspondent pas à une lignée monophylétique puisqu’elles n’ont pas d’ancêtre commun direct, mais plusieurs ancêtres, l’un pour les algues vertes et rouges, l’autre pour les algues brunes : les algues sont dites polyphylétiques.
On tente de classer le vivant en catégories hiérarchisées. L’usage est de nommer Embranchement (ou phylum) les grandes divisions. A leur tour les embranchements se subdivisent en classes, ordres et familles en sous-embranchements sous classes… Les diverses unités de la classification sont encore appelées taxons. Par exemple la classe des angiospermes se scinde en protoangiospermes, Magnoliidées, Monocotylédones et Triporées ; la sous-classe des Triporées comprend 38 ordres qui, à leur tour, se décomposent en de nombreuses familles. Afin de savoir, à chaque fois, à quel niveau de subdivision l’on se trouve, des désinences particulières ont été fixées : ainsi les ordres se terminent en ales, les familles en acées (aceae en latin) :
Ex. : l’ordre des Rosales, la famille des Rosacées (Rosaceae).
Finalement, les familles contiennent un nombre plus ou moins élevé d’espèces groupées elles-mêmes en genres. On ne définie pas, généralement, de groupement plus restreint que l’espèce. L’espèce, même si parfois on la subdivise en sous-espèces, variétés, est l’unité de base du naturaliste.
3/ Nomenclature binômiale
Chaque espèce, depuis Linné, est désignée par deux mots latins ; c’est ce que l’on appelle la nomenclature binômiale. Le premier est le nom de genre, le second le nom d’espèce.
Ex. : Rosa canina et Rosa arvensis sont deux espèces distinctes que l’on confond vulgairement sous le nom d’Églantier et qui appartiennent au genre Rosa. De plus, pour être complet, le nom d’espèce est suivi par le nom, symboliquement abrégé (par ex. : L., pour Linné), du premier botaniste qui l’a nommée selon cette nomenclature binômiale. (Avant Linné, les noms scientifiques étaient composés de plusieurs mots (nomenclature polynômiale).
4/ Notion d’espèce
On considère l’espèce comme une collection d’individus entre lesquels les différences sont faibles, alors qu’entre deux espèces, les différences sont plus ou moins profondes. C’est ce qu’on appelle le critère de ressemblance. Le verbe « ressembler » doit être pris au sens large : les caractères biochimiques, physiologiques, cytologiques, sexuels, chromosomiques entreront en considération. L’identité de ces caractères sera suffisante pour se traduire en particulier par une fécondité indéfinie des individus issus les uns des autres. C’est ce qu’on appelle le critère d’interfécondité. Au regard de la biologie moléculaire l’espèce se définit comme l’ensemble des individus susceptibles d’échanger de l’information génétique. L’espèce est définie par des caractères morphologiques qui sont seuls évidents et facile à observer.
5/ Variabilité de l’espèce, notion d’évolution
L’espèce varie ; c’est presque un de ses caractères essentielles. Dès qu’une barrière empêche l’interfécondité des individus d’une même espèce (par exemple, un isolement géographique, un isolement saisonnier, une polyploïdisation des garnitures chromosomiques), il se crée, plus ou moins rapidement, des variétés, voire des sous-espèces ou des espèces nouvelles c’est la spéciation. L’espèce est ainsi en perpétuel mouvement et, si une espèce nous paraît stable, c’est que sa vitesse d’évolution est infiniment lente par rapport à la durée de nos observateurs.
En conclusion, la notion d’espèce n’est pas un concept statique mais un concept dynamique. Pour la première fois en 1965 des caractères ont été utilisés pour retracer la phylogénie. Le critère de ressemblance porte alors sur les séquences d’aminoacides de protéines exerçant les mêmes fonctions, la comparaison des séquences homologues donne une idée de la divergence entre espèces. Cela suppose que les modifications moléculaires se font à la même vitesse pour toutes les espèces (notion « d’horloge moléculaire »).
Dans les années 90, les techniques automatiques de séquençage, tant pour les acides aminés que pour les acides nucléiques ont permis le développement de la physiologénie moléculaire.
6/ Classification artificielles, naturelles et phylogénétique
Le naturaliste va essayer de traduire cette notion d’évolution dans les classifications dites phylogénétiques. Les anciennes étaient des classifications artificielles, fondées sur des caractères faciles à observer tel le nombre d’étamines et réunissant dans les groupes communs les végétaux qui, présentent entre eux un nombre plus ou moins élevé de ressemblances profondes.
Les classifications que l’on établit maintenant tentent de rétablir l’enchainement des groupes, des plus primitifs aus plus évolués. On dira que les Gymnospermes, à ovules nus, sont plus primitives que les Angiospermes à ovules enclos, mais cela ne veut pas dire que les Angiospermes dérivent directement des Gymnospermes.
La cladistique est une méthode de classification basée sur l’analyse des caractères primitifs et évolués visant à traduire les relations phylogénétiques. Le clade est une lignée évolutive. Un cladogramme correspond à plusieurs clades successivement emboîtés. Les bifurcations (ou nœuds) correspondent à l’acquisition d’un nouveau caractère. Ces bifurcations doivent être les moins nombreuses possibles de façon à rechercher le cladogramme le plus simple. Lorsqu’un nœud possède plus de deux branches filles, la multiplication signifie que les parentés phylogénétiques précises ne sont pas résolues. La représentation dans un cladogramme des relations entre taxons sous forme dichotomique ne signifie pas que l’évolution fonctionne toujours par dichotomie. En fait la dichotomie est la seule approche possible pour comparer les relations de parentés.
Date de dernière mise à jour : 19/04/2022
Ajouter un commentaire